

Der Artikel erschien in dem mittlerweile vergriffenen UB-Heft 355 „Struktur und Funktion bei Pflanzen“ von 2010. Diese Fassung wurde an einigen Stellen erweitert, verändert bzw. aktualisiert.
Was sind Pflanzen?
Eine leichte Frage? – Aber was könnte man darauf etwa einem Außerirdischen antworten? Vielleicht: „Pflanzen sind Nichttiere.“ Also alle Lebewesen, die keine Tiere sind, sich nicht wie diese fortbewegen, fressen, mit ihren Sinnesorganen die Umwelt wahrnehmen und auf diese Umweltreize reagieren, Lebewesen die keine Nerven und Muskeln besitzen sind Pflanzen.
Wenn man das Inhaltsverzeichnis des „Strasburger, Lehrbuch der Botanik“ – seit mehr als hundert Jahren das klassische Lehrbuch der Pflanzenkunde – aufschlägt, entspricht der Inhalt dieser Vorstellung. In dem Lehrbuch werden Bakterien, Archäen, Pilze, Schleimpilze, Algen, Moose, Farne und Samenpflanzen behandelt – auch in der 37. Auflage von 2014. Einmal wird man bei genauerem Studium feststellen, dass die vorher aufgezählten „Vorurteile“ nicht alle aufrechterhalten werden können, zum zweiten sind die Unterschiede dieser Gruppen so groß, dass es nicht sehr sinnvoll erscheint, sie unter einem Oberbegriff zu fassen.
Vielleicht sollte man es deshalb mit einer positiven Definition versuchen: „Pflanzen sind grün“, sie enthalten Chlorophyll und sind damit zur Photosynthese in der Lage. Sie können aus Wasser und Kohlenstoffdioxid mithilfe von Lichtenergie und Mineralstoffen organische Stoffe wie Kohlehydrate, Lipide, Proteine und Nukleinsäuren aufbauen. Mit dieser Definition erreicht man, dass außer den Tieren auch die Pilze und die meisten Bakterien aus dem Pflanzenreich ausgeschlossen werden. Allerdings gibt es einige wirklich „echte Pflanzen“, die das Chlorophyll verloren haben und sich als Parasiten von anderen Pflanzen oder von Pilzen ihre Nährstoffe besorgen. Ihr Pflanzenstatus ist aber – wegen ihrer großen Ähnlichkeit zu verwandten grünen Pflanzen – kaum zu leugnen. Nicht sehr überzeugend an dieser Definition ist auch, dass damit sehr unterschiedliche Lebewesen, wie Blaugrüne Bakterien, viele chlorophyllhaltige Einzeller, mehrzellige, kugelige, fädige, flächige oder kompliziert strukturierte Algen sowie Moose, Farne und Samenpflanzen in einen Topf geworfen werden.
Deshalb erscheint es sinnvoller, als „Pflanzen“ Lebewesen zusammenzufassen, die nicht nur die Fähigkeit zur Photosynthese gemeinsam haben sondern auch noch weitere deutliche Ähnlichkeiten in Strukturen und Funktionen erkennen lassen. Solche Pflanzen haben einen dreidimensionalen Körper aus vielzelligen Geweben, der am Boden festgewachsen ist und sich in die Luft erhebt. Sie haben eine große Oberfläche, mit der sie im Stoffaustausch mit der Umgebung stehen und über die sie die für die Photosynthese nötige Lichtenergie auffangen können.
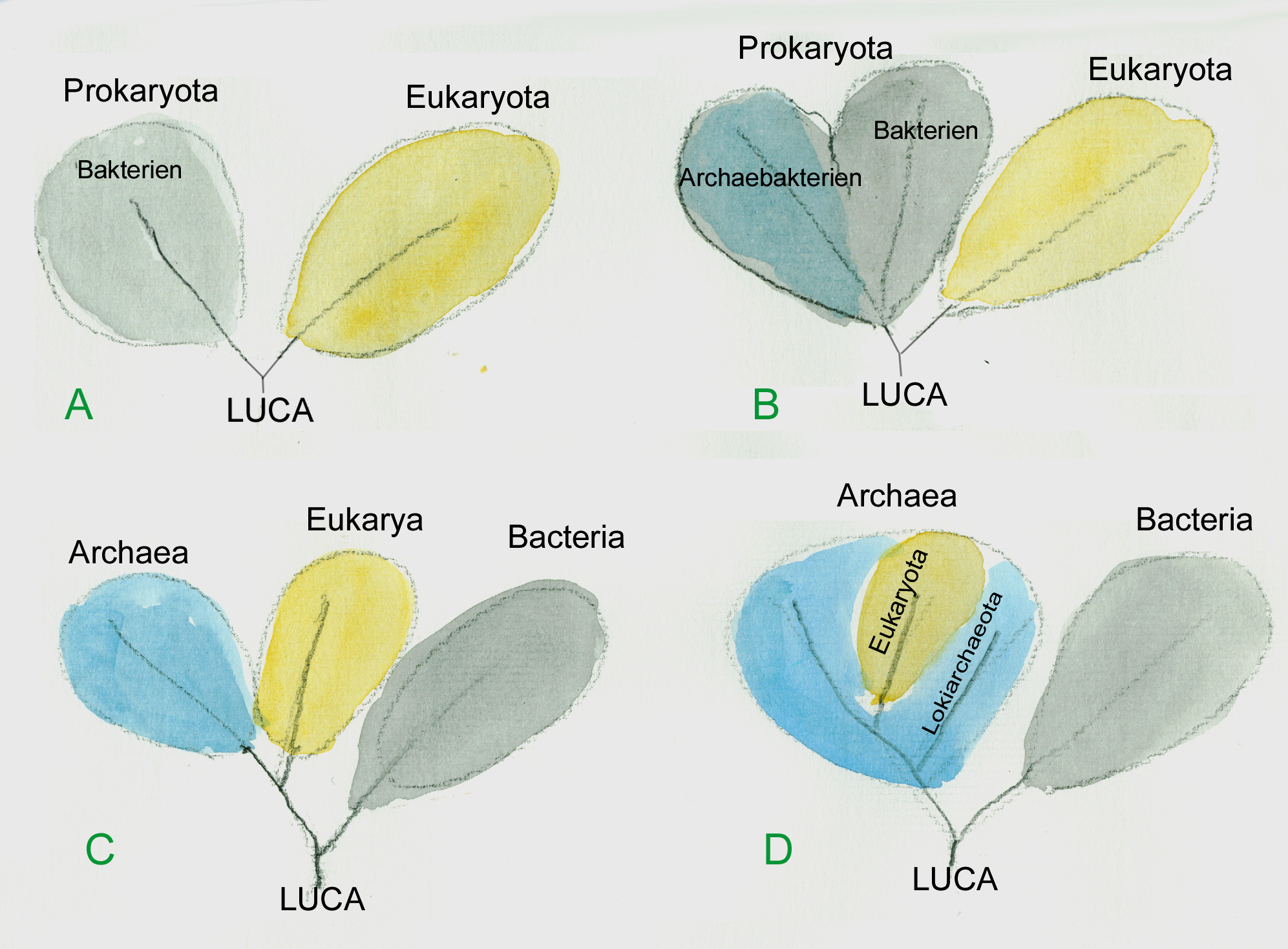
Entsprechend dem schon 1969 von Robert H. Whittaker vorgeschlagenen „Fünf-Reiche-System“ der Lebewesen werden wir uns in diesem Heft mit „Pflanzen im engeren Sinne“, also Moosen, Farnpflanzen und Samenpflanzen beschäftigen, dabei stehen die Bedecktsamigen Samenpflanzen im Mittelpunkt. Dies erscheint gerechtfertigt, da auch nach neuesten phylogenetischenn Vorstellungen ( ADL u. a. 2005) auf molekulargenetischer Basis die whittaker´schen Plantae eine monophyletische Gruppe sind.
Aus diesen Überlegungen ergibt sich eine funktionsbestimmte Definition für typische Pflanzen:
Pflanzen sind festsitzende, nachwachsende Lichtfänger mit Durchflusssystem
Festgewachsen in der Erde
Vom Lager zum Spross
Typisch für Pflanzen ist ein Vegetationskörper, der sich in die „Grundorgane“ Sprossachsen, Blätter und Wurzeln gliedert. Farnpflanzen und Samenpflanzen sind solche „Sprosspflanzen“ (Kormophyta). Die ursprünglichsten Pflanzen, die Moose, haben recht unterschiedliche Vegetationskörper, die jedoch in jedem Fall noch keine vollkommene Gliederung in Blätter, Sprossachsen und Wurzeln zeigen. Aber sie bilden wie Sprosspflanzen aus der befruchteten Eizelle einen von Schutzhüllen umgebenen Embryo, weshalb man beide zusammen auch als Embryophyta bezeichnet.
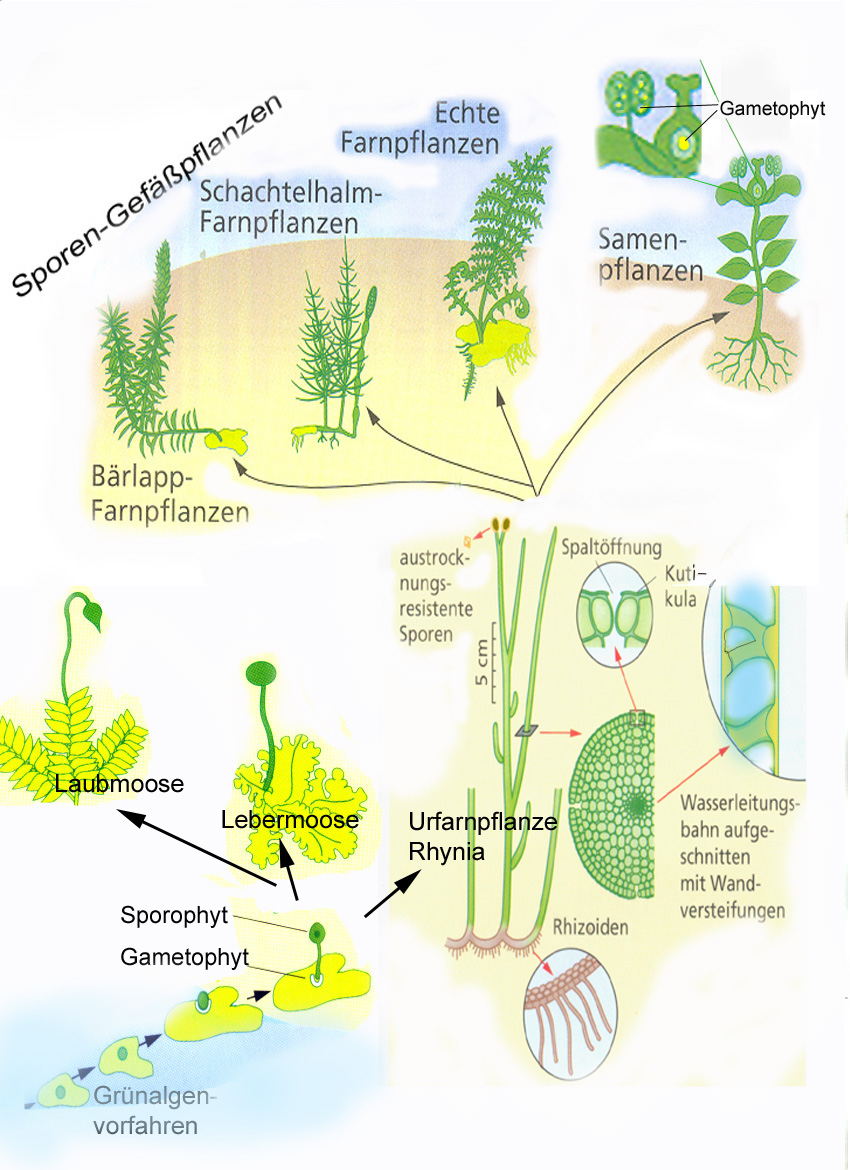
Die ersten Pflanzen, die sich deutlich in den Luftraum erhoben, entstanden vor mehr als 400 Millionen Jahren, im Silur. Diese Urfarne bestanden, wie etwa in den Gattungen Cooksonia und Rhynia, aus gabelig verzweigten Achsen, die einige Dezimeter hoch werden konnten (Bell, Hemsley 2000, Abb. 2). Bei der Differenzierung in Sprossachsen, Blätter und Wurzeln blieben die ursprünglichen Gewebe – Bildungsgewebe, Abschlussgewebe, Grundgewebe, Leitgewebe, Festigungsgewebe – erhalten. Sie sind nur entsprechend den unterschiedlichen Funktionen der verschiedenen Grundorgane jeweils anders angeordnet (Kasten Grundorgane).
Grundorgane der Sprosspflanzen

Sprossachsen
Sprossachsen ähneln in ihrem Aufbau deutlich den ursprünglichen Achsen der Urfarne. Sie dienen dazu, die Assimilations- und Transpirationsorgane, die Blätter, im Luftraum zu positionieren. Dazu benötigen sie je nach Höhe eine größere oder kleinere Biegungsstabilität, die durch besondere Festigungsgewebe und Einlagerung von Lignin in die Zellwände erreicht wird. Außerdem müssen durch Sprossenachsen Stoffe über größere Distanzen transportiert werden (Wasser- und Assimilatetransport, Transport von Mineralstoffen, Hormonen usw.). Sie wachsen über Bildungsgewebe (Meristeme) an ihren Spitzen. Zunächst ruhende Meristeme in tiefer liegenden Bereichen (Knospen) führen, wenn sie aktiviert werden, zur Ausbildung von Seitensprossen (Zweigen).
Die Sprossachsen vieler Pflanzen sind zu sekundärem Dickenwachstum in der Lage, das von einem zylinderförmigen Bildungsgewebe, dem Kambium, ausgeht.
Blätter
Laubblätter sind Organe der Photosynthese und der Transpiration. Im Gegensatz zu Sprossachsen haben sie in aller Regel ein sehr begrenztes Spitzenwachstum, das später durch basales Wachstum ergänzt wird. Sie erreichen relativ schnell ihre endgültige Größe. Sie sind im typischen Fall in Stiel und Spreite gegliedert und zeigen eine deutliche Oben/Unten-Differenzierung der Gewebe als Angepasstheit an die Ausrichtung senkrecht zum Lichteinfall. Die Ausgestaltung der Blattorgane zeigt eine sehr große Vielfalt und viele Pflanzenarten kann man an der Form ihrer Blätter erkennen. Diese morphologische Vielfalt setzt sich in bestimmtem Umfang auch noch innerhalb einer Art und sogar innerhalb eines Individuums fort. Nicht selten kommt es zu einer großen Variationsbreite zwischen Jungendblättern und Altersblättern, Sonnenblättern und Schattenblättern usw. . ).
Typische Bildungen der Blattepidemis sind die Spaltöffnungen, die aus zwei meist bohnenförmigen Zellen bestehen, die eine Öffnung umschließen, die je nach äußeren und inneren Bedingungen geöffnet und geschlossen werden kann. Sie erlauben den Pflanzen eine Regulation des Gasaustausches, insbesondere der Transpiration von Wasserdampf und der Aufnahme von Kohlenstoffdioxid. Entsprechend ihrer Funktion als Transpirations- und Photosyntheseorgane sind Blätter in der Regel dorsiventral organisiert und senkrecht zum Lichteinfall hin ausgerichtet. Die Epidermis der Blattoberseite enthält keine Spaltöffnungen, nach innen schließt an die obere Epidermis ein Gewebe aus zylinderförmigen, dicht gepackten Zellen an. Dieses Palisadenparenchym enthält besonders viele Chloroplasten. Zur Blattunterseite hin folgt das lockere Schwammparenchym, zwischen dessen rundlichen Zellen große Interzellularen ausgebildet sind, die direkt mit den Poren der Spaltöffnungen in der unteren Epidermis in Verbindung stehen und so dem effektiven Gasaustausch dienen. Von diesem Grundaufbau eines Laubblattes gibt es allerdings sehr viele Abweichungen.
Wurzeln
Während Sprossachsen und Blätter in der Regel oberhalb der Bodenoberfläche gebildet werden, sind Wurzeln die im Boden liegenden Teile der Pflanzen. Neben der Stoffaufnahme dienen sie der Verankerung der Pflanzen und damit ihrer Standfestigkeit. Entsprechen dieser Funktionen tragen sie keine Blätter und bilden auch keine Spaltöffnungen aus. Die Anordnung des mechanischen Gewebes im Zentrum stärkt die Zugfestigkeit und nicht die Biegungsstabilität. Meist handelt es sich um sehr stark verzweigte Organe, die durch diese starke Verzweigung eine große Oberfläche bilden. Diese Oberfläche wird zusätzlich an den Wurzelspitzen durch sogenannte Wurzelhaare – Auswüchse von Wurzelhaut-(Rhizodermis -)Zellen – erhöht. Nur über diese vordersten Wurzelspitzen können die Pflanzen Wasser und Mineralstoffe aufnehmen. Bei ihrem Vordringen in den Boden werden die empfindlichen Wurzelspitzen von einer Wurzelhaube (Kalyptra) aus ständig nachwachenen Zellen geschützt.
Wie die Sprossachsen haben viele Wurzeln die Fähigkeit zum sekundären Dickenwachstum.
Sekundäres Dickenwachstum
Die kontinuierliche Verdickung der Achsenorgane ist für anhaltend wachsende große Pflanzen aus zwei Gründen wichtig:
- zur Sicherung der Stabilität und
- zur Gewährleistung der Transportkapazität
Im Gegensatz zu dem primären Dickenwachstum, das von den Apikalmeristemen ausgeht, ist für das sekundäre Dickenwachstum ein zylinderförmiges Meristem im Achsenorgan, ein Kambium, verantwortlich. Es kann entweder als Restmeristem des Vegetationspunktes erhalten bleiben oder sich sekunär aus bereits differenzierten Zellen neu bilden.
Bei Einkeimblättrigen Bedecktsamern ist sekundäres Dickenwachstum selten. Die sprichwörtlich schlanken Palmenstämme sind ein Beispiel dafür, dass selbst baumförmige Pflanzen ohne sekunäres Dickenwachstum auskommen können, allerings nur mit einem sehr ausgeprägten primären Dickenwachstum. Bei den Wurzeln umgehen viele Einkeimblättrigen das Problem dadurch, dass immer wieder neue sekundäre, sprossbürtige Wurzeln gebildet werden.

Metamorphosen
Komplexe Evolutionsschritte sind nicht umkehrbar oder wiederholbar. Dies gilt jedoch nicht für den Funktionswandel. So kennt man im Pflanzenreich sehr viele Beispiele dafür, dass sich aus Sprossachsen (erneut) flache blattähnliche Organe gebildet haben und zwar in der Regel bei Pflanzen, die vorher aus anderen evolutionären Gründen ihre Blätter verloren haben. Am häufigsten kommt es zu einem solchen Blattverlust bei Pflanzen trockener Standorte (Xerophyten), die dadurch eine Verringerung der transpirierenden Oberfläche erreichen. Wenn später unter günstigeren Bedingungen der Besitz blattähnlicher Organe wieder von Vorteil wäre, kommt es jedoch nicht zu einer „Regeneration“ der verlorenen Blätter, vielmehr bilden sich aus Sprossachsenabschnitten „neue“ Blätter (Phyllokladien). Der Weg, von gefiederten Blättern wieder zu gnzrndigen Blattern zu kommen, führt über den Verlust der Blattspreite und die Verbreitrung des Blattstiels (Phyllodien, mehrfach bei der Gattung Acacia)
Besonders charakteristische Blattmetamorphosen, auf die schon Goethe aufmerksam gemacht hat, stellen die Blütenorgane dar. Unterirdische Sprossabschnitte können Wurzeln sehr ähnlich werden. Von echten Wurzeln unterscheiden sie sich jedoch oft durch kleine Schuppenblättchen und durch die Art der Verzweigung. Viele solcher unterirdischer Sprossachsen, die wie Wurzeln aussehen, sind uns recht vertraut, etwa die Rhizome (Erdsprosse) der Quecken, des Giersch oder der Winden.
Man spricht auch von „Metamorphosen“, wenn sich Grundorgane zu „neuen Organen“ umbilden, z. B. zu Dornen oder zu Ranken. Dabei sind Spross- und Blattdornen häufig, Wurzeldornen kommen nur selten vor. Dasselbe gilt für Sprossranken und Blattranken, Wurzelranken sind selten, Haftwurzeln (Efeu!) häufiger.

Emergenzen, Haare, Drüsen
Neben den Sonderstrukturen aus umgebildeten Grundorganen bilden Pflanzen auf ihren Oberflächen und auch im Inneren eine Vielfalt von Spezialstrukturen aus: Im Gegensatz zu Dornen sind Stacheln keine umgebildeten Grundorgane sondern Auswüchse oberflächennaher Gewebe. Haare entstehen aus einzelnen Oberflächenzellen, die sich jedoch mehrfch teilen können. Sie konnen rein mechanische Funktionen erfüllen, z. B. dem Verdunstungsschutz oder dem Lichtschutz dienen oder als kompakte Strukturen auch als Kletterhilfen. Besonders wirksame Fraßschutzeinrichtungen sind die Brennhaare der Brennnessel. Von manchen Haarzellen werden etherische Öle produziert (Drüsenhaare). Innere Drüsen sind oft mehrzellige Gebilde, die ihre Sekrete in interzellulare Hohlräume abscheiden. Sonderzellen in Geweben (Idioblasten) können der Stoffspeicherung oder der Ausscheidung dienen. Auch Harze und Milchsäfte können in speziellen Zellen im Inneren gebildet werden. Solche sogenannten sekundären Pflanzenstoffe sind für die Nutzung der Pflanzen als Heilkräuter oder Gewürze von Bedeutung.

Lichtfänger und Produzenten
Photoautotrophie
Das hervorragende gemeinsame Merkmal der Pflanzen ist, dass sie Lichtenergie zum Aufbau von organischen Bau- und Betriebsstoffen aus anorganischen Stoffen nutzen können (Photosynthese, vgl. z. B. UB 411, 328, 320, 249, 120, 35,). Sie fressen nicht, sie produzieren ihre Nährstoffe – Kohlenhydrate, Proteine, Lipide – selber und die Energie dazu liefert das Sonnenlicht. Dabei gelingt Pflanzen nicht nur die Assimilation von anorganischen Kohlenstoff sondern auch von Nitraten, Phosphaten und Sulfaten. Ähnlich wie Photovoltaikanlagen eine möglichst große dem Licht zugewandte Oberfläche benötigen, gilt dies auch für die „Photovoltaikanlage grüne Pflanze“, ihre Panele sind die grünen Blätter.
Diese primäre Produktion von organischen Stoffen macht sie – zusammen mit Algen und vielen Prokaryoten – zu Primärproduzenten, von denen die Stoffkreisläufe und Energieflüsse in allen Ökosystemen und in der gesamten Biosphäre ausgehen.
Dabei darf man allerdings nicht unterschlagen, dass die Pflanzen – genau wie alle anderen Lebewesen – für den lebenserhaltenden Energieumsatz Nährstoffe benötigen. Sie können diese Stoffe jedoch – im Gegensatz zu Tieren und vielen anderen Lebewesen – selbst herstellen, allerdings nur in Zellen mit Chloroplasten. Einige Pflanzen haben keine chloplastenhaltigen Zellen mehr und leben als Parasiten von anderen Pflanzen oder von Pilzen. Alle Pflanzen haben viele chloroplastenfreie Zellen und Gewebe, insbesondere ist das ganze Wurzelsystem normalerweise chloroplastenfrei und muss von den grünen oberirdischen Pflanzenteilen versorgt werden.
Primärstoffwechsel und Sekundärstoffwechsel
Wichtigstes Organell des aufbauenden (anabolen) Stoffwechsels sind die Chloroplasten, wichtigstes Organell des abbauen (katabolen) Stoffwechsels die Mitochondrien. Für die Lebensfunktionen grundsätzlich wichtige Stoffwechselwege fasst man als „Primärstoffwechsel“ zusammen. Von Zwischenprodukten (Metaboliten) des Primärstoffwechsels gehen gerade bei Pflanzen eine überaus große Anzahl von Art zu Art und von Verwandtschaftsgruppe zu Verwandtschaftsgruppe unterschiedlicher Stoffwechselwege aus, die man als Sekundärstoffwechsel bezeichnet . Bis heute kann man sich keine rechte Vorstellung von der Funktion der unübersehbaren Vielfalt dabei produzierter, meist flüchtiger organischer Substanzen machen. Ein Teil dieser Substanzen wirkt als Signalstoff, ein weiterer Teil dient der Abwehr von Fressfeinden. Zum Teil handelt es sich wohl nur um Ausscheidungen überflüssiger Stoffe. Global-ökologisch spielen solche gasförmigen Ausscheidungen von Pflanzen (VOCs von „volatile organic compounds“) eine wichtige Rolle, denn sie können sekundär organische Aerosole bilden, die die Wolkenbildung fördern (Probst 2009 in UB 349).
Stofftransport im Durchflusssystem
Voraussetzung für einen geordneten Stoffumsatz in einem Organismus ist, dass die Ausgangstoffe an den Reaktionsorten zur Verfügung stehen und dass die Endprodukte abtransportiert werden. Für die Stoffverteilung in den Pflanzen ist – anders als bei den durch ein Kreislaufsystem gekennzeichneten Tieren – der Wasserdurchfluss von den Wurzeln in die Blätter entscheidend. Auch Tiere müssen zwar ständig Wasser aufnehmen, doch im Vergleich zur im Körperkreislauf zirkulierenden Flüssigkeitsmenge ist diese Aufnahme gering – beim Menschen stehen 2-3 l täglicher Flüssigkeitsaufnahme 7000 bis 8000 l durch den Blutkreislauf bewegter Flüssigkeit gegenüber. Bei Pflanzen ist es genau umgekehrt. Auch hier gibt es in gewissem Umfang einen Kreislauf des Wassers zwischen Xylem und Phloem (Abb. 7), im Vergleich zum Wasserstrom, der vom Boden durch die Wasserleitungsbahnen bis zur Verdunstung in den Blättern führt, ist die dabei umgesetzte Menge aber sehr gering.
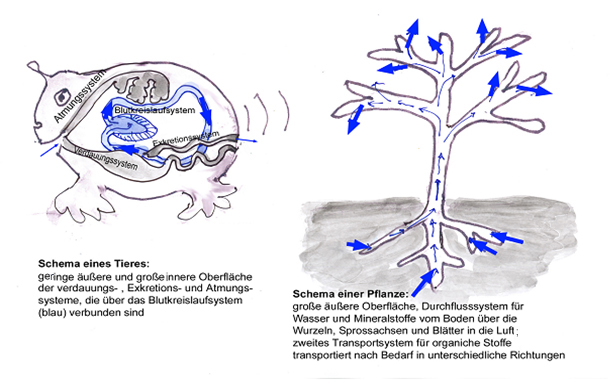
Die Notwendigkeit eines solchen Durchflusssystems hängt einmal mit der Aufnahme von Nährmineralien aus dem Boden zusammen. Für die Produktion organischer Nähr- und Baustoffe benötigen die Pflanzen außer Kohlenstoff, Wasserstoff und Sauerstoff vor allem die Elemente Stickstoff,Phosphor, Schwefel, Kalium, Calcium, Magnesium und Eisen, die als Mineralstoffionen mit der Bodenlösung aufgenommen und mit dem Wasserstrom an die Syntheseorte transportiert werden. Dies ist ein „positiver“ Grund für den Wasserdurchfluss.
Zum anderen ist die Kohlenstoffaufnahme nur über die oberirdischen Pflanzenteile aus der Luft möglich. Dabei ist das Kohlenstoffdioxid nur in sehr geringen Mengen in der Luft enthalten und um dieses wichtigste Gerüstelement aller organischen Verbindungen in ausreichender Menge aufnehmen zu können, ist eine große Oberfläche notwendig. Eine große aufnehmende Oberfläche bedeutet aber gleichzeitig eine große Oberfläche für die Wasserverdunstung. Dies ist ein „negativer“ (oder besser unvermeidlicher) Grund für den Wasserdurchfluss.
Eine große, dem Licht zugewandte Oberfläche ist – wie schon erwähnt – auch für die Photosynthese wichtig, zumal Pflanzen festgewachsen sind und dem Licht nicht hinterher laufen – höchstens ein bisschen hinterherwachsen – können. Die Verbindung mit der Wasserverdunstung ist dabei allerdings nicht ganz so eng wie bei der CO2-Aufnahme, da eine weitgehende Abdichtung der Oberflächen den Lichtfang nicht behindern würde.
Grundlage für alle Transportvorgänge in Pflanzen sind aktive oder passive Transporte durch Membranen, für Transporte über größere Entfernungen stehen spezielle Leitungsgewebe zur Verfügung.
Wasser- und Mineralstoffaufnahme in der Wurzel
Durch Diffusion bzw. Osmose wird Wasser an den Wurzelspitzen einmal in die Wurzelhaarzellen zum anderen kapillar in die Zellwände aufgenommen und osmotisch oder kapillar bis zu den Zellen der Endodermis geleitet. Dort verhindert eine Imprägnierung der seitlichen Endodermiszellwände einen weiteren kapillaren Wassertransport . Alles Wasser muss über die Membranen und das Plasma der Endodermiszellen geleitet werden. Dies gilt auch für die im Wasser gelösten Mineralstoffionen, die selektiv durch Zellmembranen aufgenommen werden, sowohl durch passiven als auch durch Stoffwechselenergie umsetzenden aktiven Transport.
Xylemtransport
Über die Endodermis gelangt das Wasser in die Leitbündel und in die bereits abgestorbenen röhrenförmigen Tracheen und Tracheiden. Treibende Kraft für den Weitertransport ist das negative Wasserpotenzial der Atmosphäre, das sich über die Spaltöffnungen, die Interzellularen der Blätter, die Kapillaren der Mesophyllzellwände und die Mesophyllzellen auf die Wassersäulen in den Leitungsbahnen auswirkt.
Dieser negative Druck kann jedoch nur zum Transport genutzt werden, wenn die Wasserfäden in den Tracheiden und Tracheen nicht abreißen. Dafür sind vor allem drei Sachverhalte entscheidend (Christian, Probst in UB 255,2010):
- Die Kohäsionskraft zwischen den Wassermolekülen
- Die Adhäsionskraft der Wassermoleküle an die Zellwände und Zellwandkapillaren
- Die Oberflächenspannung in den Zellwandkapillaren am Ende der Wassersäule
Dies würde im Prinzip ausreichen, um Wasser weit höher als 100 m zu transportieren. Da es sich jedoch nicht um reines Wasser handelt, kommt es in den Leitungsbahnen über kurz oder lang trotzdem zur Gasblasenbildung (Cavitation). Nach Kanduč et al. 2020 hängt dies mit wasserunlöslichem Lipiden in den Wasserleitungsbahnen zusammen. Werden solche Cavitäten zu groß, reißt die Wassersäule ab. Dadurch wird die Stärke der theoretisch maximal tolerierbaren Unterdrücke von -1000 bar in reinem Wasser auf weniger als -100 bar reduziert..
Phloemtransport

Auch für den Ferntransport der bei der Photosynthese produzierten Assimilate gibt es ein spezialisiertes Leitungssystem. Diese organischen Stoffe werden in wässriger Lösung über die Siebröhren bzw. Siebzellen des Phloems transportiert. Im Gegensatz zum Wasserstrom, der stets von den Wurzeln zu den Blättern fließt, kann die Assimilatelösung bedarfsabhängig in beiden Richtungen fließen. Über 90% des Siebröhrensaftes besteht aus Zuckern, v.a. aus Rohrzucker (Saccharose), außerdem sind Aminosäuren, Amide, Nucleotide (viel ATP) und organische Säuren enthalten. Motor des Transports ist der hohe osmotische Wert am Ausgangspunkt. Dadurch strömt Wasser in die Siebröhre ein. Der hydrostatische Druck führt dazu, dass Wasser am Ende der Leitung ausgepresst wird und Wasser von der Seite in die Siebröhren mit der höheren Konzentration nachfließt (Druckstrommodell). Für diesen Transport sind die siebartig durchbrochenen Querwände von Bedeutung.
Normalerweise werden die Assimilate in die wachsenden Meristeme an Spross- und Wurzelspitzen und in Speicherorgane (Knollen, Rüben) transportiert, überschüssiger Phloemsaft kann auch über besondere Drüsen (Nektarien) ausgeschieden werden. Vermutlich gehen die Nektardrüsen der Blüten auf solche Zucker-Ausscheidungsdrüsen zurück.
Blattläuse zapfen das Phloem an. Wenn sie ihre Rüssel aus den Siebröhren herausziehen, fließt der unter Druck stehende Phloemsaft noch einige Zeit nach und die überzieht die Blätter mit einem Zuckerfilm.
Durchlüftungssysteme
In den meisten pflanzlichen Gewebe schließen die Zellen nicht dicht aneinander, sie lassen vielmehr zwischen sich ein System aus Zwischenräumen (Interzellularen) frei, das letzten Endes über die oberflächlichen Spaltöffnungen mit der Außenluft in Verbindung steht. Der Stoffaustausch erfolgt über Diffusion. Dies gilt auch für Sumpf- und Wasserpflanzen und ihre unterirdischen bzw. untergetauchten Teile. Den langen Stängel der Seerosenblätter kann man als Schnorchel für das im Teichboden sitzende Rhizom auffassen.
Beim Mikrokospieren pflanzlicher Gewebe stört häufig der Luftgehalt in der Interzellularen, weil er im Nasspräparat wegen der anderen Lichtbrechung der Luft als schwarz umrandetes Objekt sichtbar wird. Deshalb ist es sinnvoll, größere Pflanzenorgane, z. B. Blätter oder Sprossachsen, bevor man sie mikroskopiert, in einer Saugflasche zu „entlüften“.
Nachwuchs
„Wachsen“ ist ein Begriff, der besonders mit Pflanzen verbunden wird, die man ja deshalb auch als „Gewächse“ bezeichnet. Wenn man Zweige abschneidet, wachsen sie nach, man kann Hecken hundertmal schneiden, Rasen tausendmal mähen und Feldhecken alle 8 Jahre „auf den Stock setzen“. Aus Knospen treiben diese gestutzten Pflanzen immer wieder neue Sprosse. Früher waren Niederwälder so ausdauernde Brenn- und Nutzholzlieferanten, nachwachsenden Rohstoffquellen. Eiben und Buchsbäume wurden von den Gärtnern der Barockgärten zu Skulpturen gestutzt.
Nachwuchs produzieren Pflanzen häufig asexuell , z. B. über Ausläufer, Ableger, Brutknospen, Brutzwiebeln. Nachwuchs kann aber auch sexuell hervorgebracht werden, bei ursprünglichen Pflanzen über einzellige Sporen, Samenpflanzen schützen ihren Nachwuchs im Samen, ähnlich wie Reptilien ihre Jungen in Eiern mit Eihüllen.
Die Entwicklung einer Pflanze lässt sich in 3 Phasen einteilen:
- embryonale Phase
- Wachstums und Reifephase
- Reproduktive Phase und Seneszenz
Bei einjährigen Pflanzen kann dieser Lebenslauf innerhalb weniger Monate oder sogar Wochen ablaufen. Das wichtigste pflanzliche Versuchsobjekt, die Acker-Schmalwand (Arabidopsis thaliana) ist ein Beispiel dafür. Bei Bäumen wird die dritte Phase oft erst nach Jahren oder sogar Jahrzehnten erreicht und kann dann nahezu unbegrenzt anhalten.
Entwicklung des Embryos

Die ersten Entwicklungsschritte bis zur Anlage der Grundorgane laufen schon im Samen ab. Die Zygote streckt sich zunächst auf die etwa dreifache Länge, dann teilt sie sich inäqual. Aus der kleineren Zelle wird der Embryo (Keimling), die größere bildet den Suspensor, der den Embryo mit dem Nährgewebe der Samenanlage verbindet. Am Ende der Embryonalentwicklung steht das Herzstadium, in dem die Keimlingsstrukturen weitgehend festgelegt sind. An den beiden Endabschnitten der apikal-basalen Achse liegen die primären Meristeme, die als selbstregulierende Stammzellensysteme alle postembryonalen Strukturen der Pflanzen hervorbringen. Dazwischen liegen die Keimwurzel, das Hypokotyl (Sprossachse zwischen Wurzel und Keimblättern) und die Keimblätter. Die weitere Entwicklung führt im Prinzip immer wieder zu denselben Differenzierungen und damit auch zu denselben Strukturen (Wurzeln, Sprossachsen, Blätter). Diese Module können allerdings – z. B. bei der Blütenbildung (.s.u.) – erheblich variiert werden. Diese Entwicklung wird von Entwicklungsgenen gesteuert und von Umweltfaktoren und endogenen Faktoren moduliert.
Entwicklungskontrolle durch Genboxen
Alle pflanzlichen Entwicklungs- und Differenzierungsvorgänge werden von Phytohormonen gesteuert. Dazu müssen zunächst die Enzyme produziert werden, die für die Hormonsynthese notwendig sind. Gleichzeitig müssen auch Gene aktiviert werden, deren Produkte für die Empfindlichkeit einer Zelle gegenüber einem Hormon zuständig sind, z. B. die Rezeptormoleküle in der Zellmembran und die Proteine für eine mögliche Signalkette im Cytoplasma. Auch für die Steuerung der Translation im Zellkern und die anschließende Aktivierung oder Hemmung der Translationsprodukte können spezielle Genprodukte notwendig sein.
Alle Entwicklungsschritte werden dadurch möglich, dass von den Genen, die in allen Körperzellen vorhanden sind, einem genauen zeitlich–räumlichen Muster folgend immer nur ganz bestimmte Gene exprimiert werden. Dieses Muster der Entwicklung wird durch eine Hierarchie von Kontrollgenen möglich, die jeweils für Transkriptionsfaktoren codieren, die andere Gene an- oder abschalten können. Für die DNA-Bindung dieser als Transkriptionsfaktoren wirkenden Genprodukte ist eine besondere Proteindömäne verantwortlich, der einem bestimmten Genabschnitt entspricht, der als „Box“ bezeichnet wird. Bei diesen Boxen handelt es sich um sehr konservative Genabschnitte, die sich über lange Evolutionsabschnitte nicht verändert haben und die zum Teil allen Eukaryoten gemeinsam sind (Seyffert 2003, S. 699 ff)
Organidentitätsgene steuern die Bildung der Blütenorgane
Eine für Pflanzen besonders bedeutende Proteindomäne ist die MADS-Box. Mittlerweile kennt man über 100 verschiedene MADS-Box-haltige Transkriptionsfaktoren, die in allen Reichen der Lebewesen vorkommen. Der Name stellt die Anfangsbuchstaben von vier Genprodukten dar, von denen zwei bei Pflanzen, eines bei der Bäckerhefe und eines beim Menschen gefunden wurden. Erste erfolgreiche Untersuchungen zur Wirkung von Kontrollgenen bei Pflanzen wurden an MADS-Box-Genen durchgeführt, die für die Entwicklung des Blütenbereiches wichtig sind und Organidentitätsgene genannt wurden.
Als Blütenorgane bezeichnet man die wirtelig angeordneten Teile einer Blüte, die von außen (bzw. unten) nach innen (bzw. oben) als Kelchblätter, Kronblätter, Staubblätter und Fruchtblätter (meist verwachsen zum Stempel) bezeichnet werden. Schon Goethe war aufgefallen, dass es bei der Blütenbildung ab und zu „Verwechslungen“ zwischen den Blütenwirteln kommen kann, dass z. B. aus Staubblättern Kronblätter werden und „gefüllte“ Blüten entstehen.
Die Entwicklung zu einer Blüte beginnt am Vegetationspunkt einer Sprossachse. Das Meristem an der Sprossspitze bildet – induziert durch äußere oder innere Faktoren – keine Laubblattanlagen mehr, sondern Anlagen von Blütenorganen. Dabei kann man drei Hierarchieebenen von Genen unterscheiden. Auf der untersten Ebene geht es um die Ausbildung der verschiedenen Blütenwirtel und dabei werden die Organidentitätsgene wirksam (Abb. 9):
Gen A wird in den beiden äußeren Wirteln exprimiert, die zu Kelch- und Kronblättern werden.
Gen B wird in den Wirteln zwei und drei exprimiert, die Kronblätter und Staubblätter bilden.
Gen C wird in den beiden inneren Wirteln exprimiert, aus denen Staubblätter und Fruchtblätter hervorgehen.
Die Wirkungsweise der Organidentitätsgene kann man sich so vorstellen, dass die Gene A, B und C jeweils nur für eine Untereinheit eines aus zwei Untereinheiten zusammengesetzten Transkriptionsfaktors codieren: A-A, A-B, B-C, C-C. Man kann in diesem Fall von einer kombinatorischen Genregulation sprechen. Die Zusammensetzung des Dimers entscheidet darüber, welche anderen Gene von dem Transkriptionsfaktor aktiviert werden. Besteht im Beispiel ein Dimer nur aus den beiden Untereinheiten A, werden Kelchblätter produziert, besteht es aus A und B, bilden sich Kronblätter aus, besteht es aus B und C, werden Staubblätter gebildet und besteht es aus zwei C´s, entstehen Fruchtblätter.
Wenn nun durch eine Mutation der Promotor, der normalerweise Gen C aktiviert, an Gen A gekoppelt wird, bilden sich keine Staub- und Fruchtblätter, sondern nur Kelch- und Kronblätter aus. Dieses Grundmuster wird durch weitere Organidentitätsgene noch etwas differenziert, außerdem gilt es nur für einen Teil der Bedecktsamer.

Licht spielt als entwicklungssteuernder Faktor bei Pflanzen eine entscheidende Rolle. Lichtabhängige Entwicklungsvorgänge (Photomorphogenese) können von Blaulicht oder Rotlicht abhängen.
Keine Vorurteile gegenüber Pflanzen !
Gängige Vorurteile sprechen Pflanzen tierliche Fähigkeiten ab: Pflanzen sind bewegungslos, sie haben keine Sinnesorgane, kein Skelett und keine Muskeln, sie haben kein Nervensystem und sie können nicht Kommunizieren und Kommunikationen verarbeiten („Denken“). Gerade bei Kindern führen solche Vorstellungen dazu, dass Pflanzen nicht unbedingt als Lebewesen eingestuft werden, deshalb sollen sie etwas ausführlicher widerlegt werden.
Bewegungslos?
Wenn man die reife Frucht eines Springkrauts anfasst, spürt man eine Reaktion zwischen den Fingern, die fast an einen Muskel erinnert. Die Frucht platzt auf, indem sich die Fruchtklappen nach innen rollen und die Samen werden ausgeschleudert. Allerdings liegen dieser Bewegung keine Muskelzellen und auch kein Nervengewebe zugrunde, denn diese spezialisierten Zellen kommen bei Pflanzen nicht vor. Trotzdem sind Reaktionen auf Umweltreize und die Verarbeitung solcher Reize möglich und in einigen Fällen führt dies auch zu auffälligen und schnellen Bewegungen.
Krümmen, Klettern, Öffnen und Schließen – Wachstumsbewegungen
Viele beobachtbare Bewegungen von Pflanzen, die auch Reaktionen auf Umweltreize darstellen können, gehen auf Wachstumserscheinungen zurück. Wenn sich in der zylinderförmigen Sprossachse einer Pflanze die wachsend Zellen auf der einen Seite stärker strecken als auf der anderen Seite, kommt es zu einer Krümmung. Wenn diese Streckungen periodisch um die Achse herum stattfinden, kommt es zu Windebewegungen. So kann das Sprossende einer Zaunwinde oder einer Bohnenpflanze kreisende Bewegungen ausführen, die wie das Suchen nach einer Unterlage aussehen. Ist diese Unterlage gefunden, wird sie relativ schnell umwunden, denn die Pflanze kann den Widerstand fühlen. Noch komplizierten wird es bei der Aufrollbewegung von Ranken. Auch hier wandert das Streckungswachstum um die Sprossachse, aber – da die Ranken am Vorder- und am Hinterende fixiert sind – muss mindestens ein Umkehrpunkt eingebaut werden. Bei der Steuerung dieser Bewegungsvorgänge von Windepflanzen spielen sowohl endogene Rhythmen als auch Umweltreize, z. B. Berührungsreize, eine wichtige Rolle.
Noch weiter verbreitet ist die Hinwendung pflanzlicher Sprossachsen zum Licht, also eine einfache Krümmungsbewegung in Richtung einer Lichtquelle. Umgekehrt zeigen Wurzeln oft eine Krümmung vom Licht weg. Solche Wachstumsbewegungen, die von einem Außenreiz ausgelöst werden, nennt man auch Tropismen und je nach Reiz spricht man von Photo-, Geo- oder Hydrotropismen, die positiv oder negativ sein können oder auch das Mittel zwischen zwei Reizeinflüssen einhalten können (Plagiotropismus), z. B. beim horizontalen Wachstum von Seitenzweigen. Nicht selten führen Pflanzenorgane richtiggehende Wachstumsprogramme durch. Die Blütenstiele von Mohnpflanzen z.B. krümmen sich vor dem Aufblühen stark ein, bei Auflühen strecken sie sich wieder. Eine ähnliche Abfolge von Krümmung und Streckung kann man bei den Blütenstände zu flach nichts beobachten.sdd
Vergleicht man solche Wachstumsbewegungen von Pflanzen mit Bewegungen von Tieren, so ergeben sich deutliche Unterschiede. Insbesondere sind diese auf Wachstumsvorgänge beruhende Bewegungen alle sehr langsam, sie können von uns nur indirekt wahrgenommen werden und sie sind eher mit den Form- und Proportionsveränderungen vergleichbar, die im Laufe der Keimesentwicklung von Tieren auftreten.
Explodieren, Schleudern, Klappen… – Turgorbewegungen
Die zweite große Gruppe pflanzlicher Bewegungen beruht auf Änderungen des Zellbinnendrucks von Pflanzenzellen, des sogenannten Zellturgors, der in der Regel durch den unterschiedlichen Wassergehalt der Zentralvakuolen der Pflanzenzellen zustande kommt. Dieser wird wiederum über den osmotischen Wert gesteuert.
Für fast alle Pflanzengruppen sind Spaltöffnungen charakteristisch. Das sind von zwei Schließzellen umgebene Poren in der Außenhaut (Epidermis), die in Abhängigkeit von äußeren und inneren Reizen geöffnet und geschlossen werden können. Ist der Turgor der bohnenförmigen Schließzellen hoch, bilden sie eine nahezu kreisförmige Gestalt, die in der Mitte einen Porus offen lässt. Ist der Turgor niedriger, sorgt der Druck des übrigen Gewebes dafür, dass die Schließzellen aneinandergepresst werden. Dies ist die klassische Bewegungsreaktion, es gibt viele Abwandlungen und Sonderformen, die durch die besondere Gestalt der Schließzellen und ihre Anordnung in der Epidermis zustande kommen. Turgorschwankungen sind aber in jedem Fall der Motor der Bewegung. Ursache für diese Turgorschwankungen sind Schwankungen im Ionengehalt, insbesondere im Kaliumionengehalt der Schließzellvakuolen und dieser Ionengehalt kann durch aktive Pumpmechanismen verändert werden. Die Regulation der Spaltöffnungsbewegung sorgt dafür, dass die Pflanzen mit der schwierigen Aufgabe zurecht kommen, in der trockenen, kohlenstoffdioxidarmen Atmosphäre weder zu verhungern noch zu verdursten.
Spektakulärer, da ohne optische Hilfsmittel sichtbar, sind andere Turgorbewegungen von Pflanzen. Da wären so auffällige Beispiele wie die „Schamhafte Mimose“ oder Sinnpflanze (Mimosa pudica) oder die Venusfliegenfalle (Dionaea muscipula) zu nennen. Die Reizreaktionen dieser Pflanzen erinnern wirklich an nerven- und muskelgesteuerte Bewegungen von Tieren. Für die Bewegungen sind nicht nur einzelne Zellen, wie bei den Spaltöffnungen, sondern ganze Gewebe verantwortlich, deren Turgor aktiv verändert werden kann. Dabei kommt die schnelle Bewegung in der Regel durch einen plötzlichen Turgorabfall zustande, die anschließende „Regeneration“, die mit einem Aufbau des alten Turgorzustanden verbunden ist, dauert etwas länger.
Ähnlich wie das Öffnen und Schließen einer Tür werden Turgorbewegungen zwar oft von Außenreizen bewirkt, sie laufen aber unabhängig von der Richtung der Reizquelle ab. Solche Bewegungen werden als Nastien bezeichnet
Neben reversiblen Turgorbewegungen kennt man irreversible. Hierzu zählt das oben angesprochene Beispiel des Springkrauts. In den schmalen zylinderförmigen Zellen der äußeren Fruchtwand wird ein hoher Turgordruck aufgebaut, der schließlich zu einem Aufreißen der Frucht an präformierten Nähten zwischen den Fruchtblättern führt. Sind die Fruchtblätter erst voneinander getrennt, können sich die zylinderförmigen Zellen ausdehnen und dadurch werden die Fruchtblätter nach Innen eingerollt. Dabei spielt allerdings auch noch die elastische Zellwand eine Rolle. Wenn sich die Zellen der Außenwand erst einmal abgerundet haben, ist eine Rückkrümmung er Fruchtklappen nicht mehr.
Verbiegen und Verdrehen – Quellungsbewegungen
Für das Funktionieren von Pflanzen sind auch Bewegungen von Bedeutung die keine physiologischen sondern rein physikalische Ursachen haben, hierzu zählen Quellungs- und Kohäsionsbewegungen. Entscheidend für diese Bewegungen sind Zellulosestrukturen (Abb. 3). Zellulose ist aus fädigen Molekülen aufgebaut, die zu sogenannten Mizellen zusammengefasst sind. Diese Zellulosesemizellen können in unterschiedlicher Richtung in eine Zellwand eingelagert sein, ähnlich wie die Faserstrukturen im Papier. Zwischen die Mizellen kann Wasser eingelagert werden. Dies führt zu einer Ausdehnung und zwar stärker quer zur Faserrichtung als in Faserrichtung. Sind die Faserrichtungen in zellulosehaltigen Pflanzenstrukturen geordnet aber in verschieenen Schichten unterschiedlich, so kann dies beim Quellen zu Krümmungsbewegungen führen. Bei den Schuppen von Nadelholzzapfen zum Beispiel, aber auch bei vielen Fruchtklappen oder -zähnchen sind die Zellulosemizellen außen vorwiegend quer und innen längs angeordnet. Die stärkere Außenquellung bei Wassereinlagerung führt dazu, dass sich Nadelholzzapfen bei Feuchtigkeit schließen und bei Trockenheit durch nach außen Krümmen der Schuppen wieder öffnen. Dadurch wird erreicht, dass die Samen nur bei Trockenheit ausfallen können, was für deren Windverbreitung von Vorteil ist. Weitere auffällige Beispiele für solche Quellungsbewegungen zeigen viele Öffnungsklappen von Früchten oder die Peristomzähnchen an den Kapseln von Moossporogonen.
Je nach Anordnung der Micellen kann bei diesen Bewegungen nicht nur zu Krümmungen sondern auch zu Schraubungen kommen. Besonders einrucksvoll ist die lange, wie ein Drillbohrer wirkene Fruchtklappe des Reiherschnabels.
Auch solche Quellungsbewegungen können mit dem Bruch von präformierten Nähten verbunden sein. Bei vielen Schmetterlingsblütlern öffnen sich die Hülsen nahezu explosionsartig, beim Blauregen (Wisteria sinensis) zum Beispiel mit einem richtigen Knall. Dies kommt dadurch zustande, dass bei Austrocknung sich allmählich eine Spannung aufbaut, die schließlich zum Aufreißen an präformierten Nahtstellen führt. Oft kann man dieses Aufreißen als kleinen Knall wahrnehmen (z. B. bei Lupinen, beim Blauregen oder bei der Blatterbsen-Wolfsmilch).
Bei dem Aufreißen von Farnsporangien, spielt die Köhäsion der Wassermoleküle eine wichtige Rolle.
Von einem Ort zum andern
Pflanzen können nicht aktiv kriechen, laufen, schwimmen oder fliegen, sich zum Beispiel gerichtet auf eine Reizquelle (Taxie) zubewegen oder einer Spur folgen. Passive Ortsbewegungen sind aber möglich. So sind viele Wasserpflanzen nicht festgewachsen. Sie fluten frei an der Oberfläche oder untergetaucht im Wasserkörper und werden von Strömungen mitgetragen. Nur so konnte sich die Wasserhyazinthe (Eichhornia crassipes) – ursprünglich im Amazonasgebiet zuhause – über fast alle tropischen und subtropischen Gewässer verbreiten und die Kanadische Wasserpest (Elodea canadensis) hätte andres nicht in kurzer Zeit zu einer die Schiffahrt bedrohenden „Pest“ in europäischen Gewässern werden können.
Einjährige Wüsten-oder Steppenpflanzen können, wenn sie nach der Samenreife absterben, zu „Wüstenrollern“ werden, die vom Wind beträchtliche Strecken über den Steppenboden gerollt weren und dabei die Samen allmählich ausstreuen.
Dies schafft den Übergang zu den Pflanzenteilen, die auf Fortbewegung spezialisiert sind, weil sie der Ausbreitung dienen sollen. Sie werden Diasporen genannt. Diasporen können Sporen, Samen, Früchte, Teilfrüchte und vegetative Ausbreitungseinheiten wie Brutknospen, Brutzwiebeln, Spross- und Wurzelknollen oder Butkörper sein. Triebkräfte bzw.Transportmittel für die Ortsbewegung sind Wind, Wasserströmung, die Schwerkraft, Tiere und der Mensch. Dabei wird in der botanischen Literatur streng zwichen Ausbreitung, dem Fortbewegungsvorgang einer Diaspore, und der oft daraus resultierenden geografischen Verbreitung einer Pflanzenart unterschieden.
Knochenlos?
Die Bewegungen der Tiere kommten i. A. durch das Zusammenspiel von Muskeln und Skelett zustande. Dabei kann es sich um ein inneres Knochenskelett (Wirbeltiere), einen äußeren Skelettpanzer (Indsekten und andere Panzerhäuter – Ecdysozoa) oder auch ein Hydroskelett (Regenwurm und andere Würmer) handeln. Pflanzen haben sowohl ein den Knochen entsprechendes Innenskelett, das aus Geweben mit verdickten Zellwänden besteht, als auch ein Hydroskelett.
Tierzellen haben keine Zellwände, die harten Skelettelemente bilden sich aus Abscheidungen der Zellen in den extrazellulären Raum. Demgegenüber besitzt jede Pflanzenzelle eine feste Zellwand, die vor allem aus langkettigen Polysacchariden, insbesondere aus Zellulose, besteht. Es können aber weitere Stoffe eingelagert oder aufgelagert sein, z. B. Lignin bei verholzten Zellwänden oder Siliziumdioxid bei Gräsern und Schachtelhalmen. Pflanzenzellen machen in der Regel nach ihrer Bildung eine lange Phase der Volumenzunahme durch, wobei eine starre Zellwand sehr hinderlich wäre. Deshalb sind die Polysaccharidfasern in den Zellwänden junger Pflanzenzellen noch wenig fixiert und der Volumenzunahme folgend können immer neue Fasern eingebaut werden. Erst wenn die Zelle ihre endgültige Größe erreicht hat, kommt es zu einer zunehmenden Stabilisierung und Verfestigung der Wände, die vor allem durch einen regelmäßigen Wechsel in der Textur der Zellulosefibrillen und zunehmenen Querverbindungen zwischen den Fibrillen erreicht wird.
Durch die Bildung von Zellen mit sehr dicken Zellwänden können mechanisch stark belastbare Gewebe entstehen. Dabei kann es sich um lange Zellfasern handeln, die vor allem Zug- und Dehnungsstabilität bewirken und die von der Textilindustrie auch als „Pflanzenfasern“ genutzt werden (Baumwolle, Hanf, Lein, Jute, Sisal usw.) oder um isodiametrische Steinzellen, die vor allem Druckbelastungen standhalten. Durch die kompakte Anordnung von Zellen mit stark verdickten und verholzten Zellwänden kann eine hohe Druck- und Biegungsstabilität erreicht werden.
Besonders eindrucksvoll sind die kompakten Holzkörper der großen Bäume, die allerdings zum großen Teil aus abgestorbenen Zellen bestehen. Der lebende Baum bildet nur einen hautartigen Überzug über dem toten Holzskelett. Es gibt aber auch beeindruckene Beispiele für Leichtbauweisen. Viele Pflanzenkonstruktionen zeigen, wie mit möglichst geringem Materialaufwand möglichst große Stabilität oder Festigkeit erzeugt werden kann, z. B. reißfeste Blätter, biegungsstabile Grashalme, oder tragfähige Säulenkakteen.
Typisch für Pflanzenzellen sind außerdem Vakuolen mit wässerigem Inhalt, die bei ausgewachsenen Zellen meist den größten Anteil des Zellinneren ausmachen (Zentralvakuole). Je höher der Gehalt der Vakuole an osmotisch wirksamen Substanzen, je niedriger also ihr Wasserpotenzial, desto mehr Wasser wird von außen angezogen und desto größer wird der Binnendruck des Zellkörpers gegen die Zellwand, den man auch als Turgor bezeichnet. Für die Stabilität eines Pflanzensprosses ist dieses durch den Turgor gebildete Hyroskelett von großer Bedeutung. Dies kann man daran erkennen, dass Pflanzen bei Wasserverlust „verwelken“. Sie sind dann nicht mehr in der Lage, ihr eigenes Gewicht zu tragen: Ihre Blätter hängen schlaff herab und sie biegen sich zum Boden.
Vakuolen dienen aber auch als Stoffreservoir für die Pflanzenzellen. Hier können organische Säuren gespeichert werden, ebenso wasserlösliche Farbstoffe. Es kann auch zum Ausfallen schwer löslicher Verbindungen in Vakuolen kommen, die dann mit dem Absterben der entsprechenden Pflanzenteile, z. B. der Blätter, auf diese Weise ausgeschieden werden. Eine wichtige Funktion der Vakuolen ist auch die der Wasserspeicherung. Besonders große Vakuolen kommen in den Früchten vor und wir genießen den leckeren Vakuoleninhalt, wenn wird die saftigen Früchte essen oder die ausgepressten Fruchtsäfte trinken.
Sinnenlos?
Pflanzen haben keine richtigen Sinnesorgane wie Augen und Ohren, sie sind aber durchaus in der Lager, physikalische und chemische Umweltreize wahrzunehmen, insbesonere Licht und Schatten, Schwerkraft, Erschütterungen und Berührungen, Wärme (bzw. Temperaturunterschiede) sowie Wasser und bestimmte Mineralstoffe.
Für die Lichtwahrnehmung sind v.a.zwei Pigmentsysteme, das Phytochromsystem und das Phototropinsystem verantwortlich. Das Phytochrom ist – wie das Chlorophyll und das Häm – ein Tetrapyrol, das allerdings keinen Porphyrinring bildet, sondern in offenkettiger Form vorliegt. Durch die Absorption von Photonen kann es seine Konfiguration ändern. Dadurch wirkt es wie ein lichtabhängiger Schalter. Bei Belichtung mit hellrotem Licht der Wellenlänge 665 nm wird Phytochrom in ein Molekül umgewandelt, das dunkelrotes Licht (735 nm) absorbiert, bei Belichtung mit dunkelrotem Licht wird dieses wieder in das Hellrot-absorbierende Phytochrom zurück verwandelt. Die jeweils zuletzt eingestrahlte Lichtqualität entscheidet über die ausgelöste Entwicklung. In der Zelle ist das Phytochrom an ein Protein gebunden. Das Phytochromsystem ist z. B. für die Samenkeimung von Licht- und Dunkelkeimern und für das extreme Streckungswachstum im Dunkeln (Etiolement, Vergeilung) verantwortlich.
Für die Krümmung von Pflanzensprossen zum Licht (Photropismus) ist nur der blaue Anteil des Lichts verantwortlich. Als wahrnehmendes Pigmentsystem konnte das Chromoprotein Phototropin nachgewiesen werden. Blaulicht wird von dem Apoprotein es Phototropins, einem Flavinmononucleotid, absorbiert und dies bewirkt, dass ein Phosphat an das Phototropin angehängt wird. Das so aktivierte Phototropin setzt eine Signaltransduktionskette in Gang, die dazu führt, dass im Sprossmeristem verstärkt das Phytohormon Auxin produziert wird. Es wird zunächst auf die dunkle Seite des Vegatationskegels verlagert und dann vorwiegend über die basalen Zellmembranen weitergegeben. Dafür sind bestimmte Effluxproteine und Afflux-Carrier verantworlich. Das Auxin verstärkt das Streckungswachstum der Zellen dadurch, dass die H+-ATP-aseaktivität und die K+-Aufnahme durch die Zellmembran verstärkt werden.

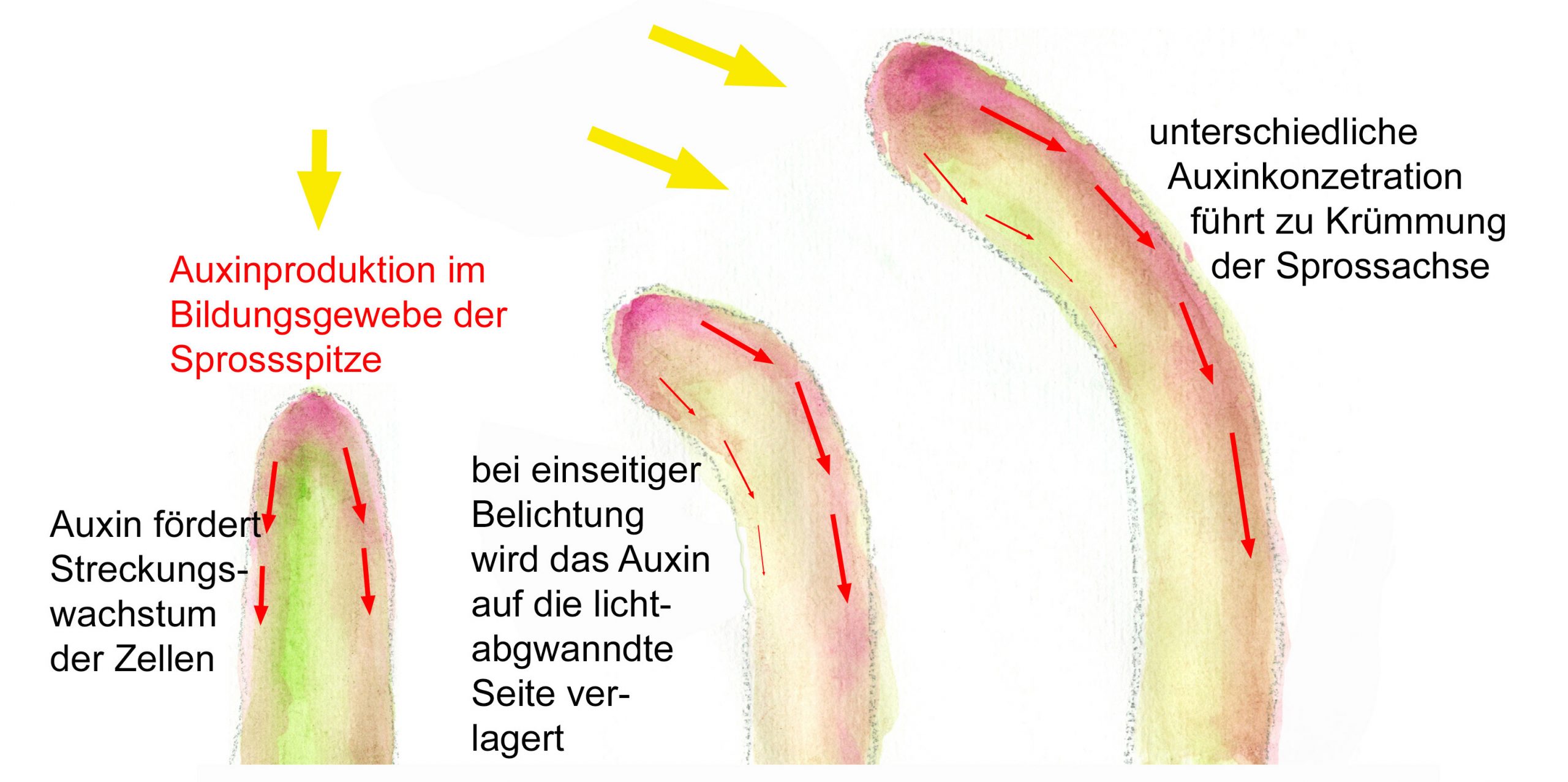
Auch zur Wahrnehmung der Schwerkraft bzw. der Massenbeschleunigung haben Pflanzen sehr empfindliche Einrichtungen, die in mancher Hinsicht an Schweresinnesorgane von Tieren erinnern. Da in Schwerkraft-empfindlichen Pflanzenteilen wie Wurzelhauben, Keimscheiden und Sprossachsen auffällig assymmetrisch gelagerte Stärkekörner (in Amyloplasten) auftreten, spricht man auch von Statolithenstärke. Es wir vermutet, dass der entscheidende Reiz die partielle Dehnung der äußeren Zellmembran ist, die durch das Gewicht des Protoplasten ausgelöst wird. Die Statolithenstärke erhöht das Gewicht.
Viele Pflanzen – v.a. kletterne Sprossachsen und Ranken – können Berührungsreize wahrnehmen. Bei der fleichfessenen Venus-Fliegenfalle gibt es eine Reizwahrnehmung durch Sinnesborsten, ein Aktionspotenzial und eine Erregungsleitung . Noch ausgeprägter ist die Erregungsleitung bei der Sinnpflanze (Mimosa pudica).
Wurzelspitzen können chemische Reize wahrnehmen, z. B. Phosphationen, Wassermoleküle und Sauerstoffmoleküle. Auf diese Weise können Baumwurzeln kleinste Risse in unterirdischen Wasserleitungen aufspüren und in den Wasserleitungen dann verstopfende „Wurzelzöpfe“ bilden.
Sprachlos und gedankenlos?
Vor mehr als 35 Jahren erregte ein Buch mit dem Titel „Das geheime Leben der Pflanzen“ viel Aufsehen (Tompkins, Bird 1973). Die Autoren stellen darin „Pflanzen als Lebewesen mit Charakter und Seele und ihren Reaktionen in physischen und emotionalen Beziehungen zum Menschen“ dar. Sie schildern, wie eine ausgedehnte sprachliche Kommunikation mit Pflanzen möglich sei und wie man die Gefühle seines Gummibaumes oder seiner Palmlinie durch Zuspruch günstig oder ungünstig beeinflussen könne. Pflanzen werden in diesem Buch gewissermaßen als „bessere Menschen“ dargestellt. Von Botanikern und Pflanzenphysiologen wurde diese Veröffentlichung natürlich nicht ernst genommen und zu Recht ist es schnell still geworden um die angeblich objektiven Experimente der an Lügendetektoren und Oszilloskope angeschlossenen Pflanzen. Allerdings weiß man heute viel mehr über Signalaufnahme, Verarbeitung und Weitergabe von Pflanzen und über Formen pflanzlicher Kommunikation als vor 30 oder 40 Jahren und dabei wurden durchaus erstaunliche Fähigkeiten entdeckt. So besitzen Pflanzen ein großes Repertoire an Signalstoffen, die sie in Abhängigkeit von Umwelteinflüssen einsetzen können, um im eigenen Pflanzenkörper oder auch bei Nachbarpflanzen und sogar bei Tieren Reaktionen hervorzurufen.
Signalstoffe, die steuernd und regelnd in die inneren Entwicklungs- und Stoffwechselprozesse einer Pflanze eingreifen, werden in Analogie zu tierlichen Hormonen als Phytohormone bezeichnet. Phytohormone sind relativ kleine Moleküle, die Informationen von ihrem Bildungsort zu Zielzellen und Zielgeweben übertragen. In der Regel setzen sie in den Zielzellen eine Signaltransduktionskette in Gang, die zu einer Reaktion der Zelle führt . Diese Reaktion kann direkt im Cytoplasma stattfinden, sie kann aber auch über Transkriptionsfaktoren zum An- oder Abschalten von Genen führen. Dabei spielen Rezeptoren in der Zellmembran, verschiedene Proteine (G-Proteine, Proteinkinasen, phosphorylierte Proteine als Transkriptionsfaktoren) und kleine, als sog. sekundäre Botschafter (second messenger) wirkende Moleküle und Ionen wie Ca++, und cyclisches Adenin- bzw. Guanin-Monophosphat (cAMP, cGMP) eine Rolle (Tab. Phytohormone).
Viele Pflanzen produzieren chemische Abwehrstoffe, mit denen Fressfeinde oder Krankheitserreger abgewehrt werden können. Oft werden solche Stoffe erst produziert, wenn die Pflanzen von pflanzenfressenden Insekten oder anderen Herbivoren angeknabbert werden. Die Konzentrationen von Limonen und anderen Monoterpenen im Gewebe von Fichten und Lärchen zum Beispiel erhöhen sich als Reaktion auf Verletzungen. Limonen hat eine starke toxische Wirkung auf Borkenkäfer. Auch das Alkaloid Nikotin, das z. B. in Tabakpflanzen vorkommt, ist ein hochwirksamen Insektengift. In beschädigten Blättern erreicht es eine bis zu 10fach höhere Konzentration als in unbeschädigten.
Bestimmte durch Fraßfeinde hervorgerufene Verletzungen können Pflanzen dazu veranlassen, Stoffe wie Jasmonate als Signalstoffe an die Umgebung abzugeben. Diese Pheromone regen dann auch bei Nachbarpflanzen die Produktion von Abwehrstoffen an. Die Pheromonwirkung ist nicht nur auf Pflanzen derselben Art beschränkt. Manche Pflanzenarten setzen als Reaktion auf Herbivorenbefall sogar Substanzen frei, die die Parasiten dieser Pflanzenfresser anlocken. Maispflanzen, die durch den Fraß von Raupen der Zuckerrübeneule beschädigt wurden, produzierten unterschiedlich flüchtige Terpenoide, die nachweislich als Lockstoff für die auf den Raupen parasitierende Schlupfwespen (Cortesia marginiventris) wirken. . Die Signalketten, die von der Vielfalt flüchtiger Pflanzensubstanzen an ihren Zielen ausgelöst werden können, sind bisher zum großen Teil unbekannt, aber es gibt auch schon viele erforschte Beispiele (Abb. 10).
Besonders vielfältig ist der Signalaustausch der Pflanzen mit ihren Bestäubern, sowohl über optische als auch über chemische Signale

Ebenso wie bei der tierlichen Kommunikation spielt auch bei Pflanzen die Täuschung eine Rolle (Kasten Duftmimikry bei Orchideen).
Interesse wecken für das Andere? – Pflanzenkunde unterrichten
Pflanzen sind für Kinder und Jugendliche i.d.R. weniger faszinierend als Tiere. Dies belegen mehrere empirische Studien (z. B. Hesse 2000). Während schon Kleinkinder von Tieren begeistert sind und ihnen ihre ganze Aufmerksamkeit zuwenden, sind Pflanzen selbst für Kinder im Grundschulalter in der Regel gar keine richtigen Lebewesen. Pflanzen, Nicht-Tiere, reagieren nicht, wenn man sie anspricht oder streichelt, sie bewegen sich nicht von der Stelle, sie bellen, brüllen oder zwitschern nicht und sie lassen keinen Schmerz erkennen. Die große Bedeutung, die Pflanzen für das Landschaftsbild haben, ist für Kinder ebenfalls nicht wichtig, da sich Interesse und Gefühle für Landschaftsästhetik erst später entwickeln. Allerdings gibt es durchaus animierende Gefühle zu Pflanzen, die mit Ästhetik, Abenteuerlust oder Freude an technischen Funktionen zu tun haben:
- Aus Pflanzen etwas gestalten: Blumenstrauß-Pflücken, Kränze und Blumenketten binden…
- Aus Pflanzenteilen etwas konstruieren: Weidenpfeifen und Panflöten bauen, Wasser durch Löwenzahnstängel leiten, aus einem Schilfblatt ein Segelschiffchen bauen, einen Haselzweig zum Flitzebogen machen…
- Pflanzenumgebungen für Abenteuer nutzen: auf Bäume klettern und Baumhäuser bauen, an einer Liane schaukeln, Baumstämme als Wippe nutzen, im Maisfeld Verstecken spielen, sich in Laubhaufen vergraben…
Auch das Phänomen der Samenkeimung und des Wachstums oder das Ableger Großziehen kann Kinder und Jugendliche faszinieren.
Pflanzenkunde-Unterricht muss an solche Erfahrungen und Vorstellungen anknüpfen. Entsprechende Versuche hat es in der Biologiedidaktik immer wieder gegeben: UB 184 „Kreative Botanik“, UB 275 „Pflanzen züchten und vermehren“, UB 286 „Außergewöhnliche Pflanzen“, „Pflanzen stellen sich vor“, „Gärten zum Leben und Lernen“ usw.
Im Rahmen des Modellversuches „Praxis integrierter naturwissenschaftlicher Grundbildung“ (PING) des Landes Schleswig-Holstein wurde für den 5/6. Jahrgang eine UE „Ich und die Pflanzen“ entwickelt, bei der der lebensweltliche Bezug der einzelnen Themen besonders wichtig genommen wird. Dabei wird allerdings in Kauf genommen, dass es oft nicht eigentlich um „Pflanzenkunde“ sondern um Selbsterfahrung („Phantasiereise zur blauen Blume“) oder um irgendwelche Nutzungen von Pflanzen oder pflanzlichen Produkten geht („Wie energiesparend kann ich mit Holz kochen“). Dabei ist der Ansatz sicherlich richtig, bei der Pflanzenkunde verstärkt von eigenen Erfahrungen oder Erfahrungen mit Tieren auszugehen, z. B. durch Fragestellungen wie „Können Pflanzen sehen?“, „Können Pflanzen atmen?“, „Können Pflanzen fühlen?“, „Können Pflanzen laufen, schwimmen, fliegen, klettern…?“ oder „Können Pflanzen um Hilfe rufen?“ . Dabei könnte von der eigenen Erfahrung (Wie klettere ich? Welche Hilfsmittel nutze ich zum Klettern?) die Funktion des Kletterns bei Pflanzen und dann der Bau (Pflanzenlösung des Funktionsproblems) behandelt werden. Ästhetische Erlebnisse mit schönen Blüten oder bizarren Blattmustern können der Ausgangspunkt für Fragen nach der biologischen Funktion und der physikalischen Grundlage der Phänomene sein – z. B. leuchtende Blütenfarben durch Totalreflexion an Interzellularen, Fettglanz durch Reflexion an Stärkeschichten, Samteffekt durch Lichtabsorbtion an feinen Papillen usw.
Gerade weil Kinder Pflanzen zunächst als keine echten Lebewesen oder Mitgeschöpfe ansehen, ist der Überraschungseffekt groß, wenn sie an Pflanzen tierliche Fähigkeiten entdecken können, z. B. rasche Bewegungsreaktionen auf Reize oder das „Fleischfressen“. Dabei kann sich die Lehrkraft Charles Darwin zum Vorbild nehmen. In seiner Autobiografie schreibt er: „Es hat mir immer große Freude bereitet, die Pflanzen in der Stufenleiter organisierter Wesen zu erhöhen. Ich empfand daher ein besonderes Vergnügen, als ich zeigen konnte, wie viele und wie wunderbar schön angepasste Bewegungen die Spitze einer Wurzel besitzt“ (Darwin, Erinnerungen…1876-1881, Aulis,Köln 1982, S.159).
Für unmittelbare Erfahrungen, Beobachtungen und Experimente am lebenden Objekt sind Pflanzen besonders gut geeignet., da die Untersuchungsobjekte i. d. R. leicht zu beschaffen sind , und auch, weil ethische Probleme, wie sie beim Experimentieren mit Tieren auftreten können, hier keine Rolle spielen. Schließlich stehen Pflanzen beim „Biologieunterricht im Freien“, bei Geländepraktika, Exkursionen, Schulgartenprojekten und Klassenfahrten schon deshalb im Vordergrund, weil sie die Struktur einer Lebensgemeinschaft maßgebend prägen, weil sie als leicht zugängliche Zeigerorganismen dienen können und weil man über Pflanzenarten viele ökologische Zusammenhänge erschließen kann. Auch wenn bei der Freilandbiologie ökologische Fragestellungen im Vordergrund stehen, kann man gerade auf Exkursionen auch viele Form- und Funktionszusammenhänge beobachten, erleben und verstehen.
Der richtige Weg einer zwar „humanzentrierten Pflanzenkunde“, der aber dazu führt, dass die Lernenden auch die Pflanze selbst und nicht nur ihre Beziehung zu derselben als interessant begreifen, bleibt das schwer erreichbare aber lohnende Ziel.
Quellen
Adl, S., M. u. a.( 2005) :The New Higher Level Classification of Eukaryotes with Emphasis on the Taxonomy of Protists. J. Eukaryot. Microbiol., 52(5), 2005 pp. 399–451 http://myweb.dal.ca/asimpso2/sopclass.htm
Bell, A.D. (1994): Illustrierte Morphologie der Blütenpflanzen, 335 S.. Stuttgart: Ulmer
Bell, P.R., Hemsley, A.R. (20002): Green plants. Their origin and diversity. Cambridge Univ. Press
Bildungsserver Hessen http://lernarchiv.bildung.hessen.de/sek_i/biologie/themen/botanik/index.html
Braune, W., Leman A., Taubert, H. (20028): Pflanzenanatomisches Praktikum I. Spektrum, Heidelberg/Berlin: Spektrum
Darwin, C. (1982): Erinnerungen an die Entwicklung meines Geistes und Charakters…Urania Leipzig 1982, Lizenzausgabe; Köln: Aulis
Frey, W., Lösch, R. (20103): Lehrbuch der Geobotanik. München: Elsevier
Goethe, J.-W. von: Die Metamorphose der Pflanzen http://www.zeno.org/Literatur/M/Goethe,+Johann+Wolfgang/Naturwissenschaftliche+Schriften
Heldt, H.-W (20033).: Pflanzenbiochemie. Heidelberg/Berlin: Spektrum
Hesse, M. (2000): Erinnerungen an die Schulzeit – Ein Rückblick auf den erlebten Biologieunterricht junger Erwachsener. Zeitschrift für Didaktik der Naturwissenschaften 6, 187-201
Institut für Qualitätssicherung Schleswig-Holstein (IQSH): PING–Material (2006):
Ich und die Pflanze http://ping.lernnetz.de/pages/n350_DE.html
Kanduč, M. et al. (2020): Cavitation in lipid bilayers poses strict negativ pressure tability limit in biological liquids. PNAS, vol.117,pp.1033-1039
Lüttge, U., Kluge, M., Thiel, G. (2010): Botanik. Weinheim: Wiley-VCH
Martin, K. (2002): Ökologie der Biozönosen. Berlin/Heidelberg: Springer
Munk, K. Hrsg (2009): Botanik. Taschenlehrbuch Biologie, 573 S., Stuttgart: Thieme
Nabors, M. W. (2007): Botanik. München:. Pearson Deutschland
Preitschopf, W.: Sekundärstoffe http://infowis.de/kapitel/sekundae.html
Probst, W. (1999): Halme und Stängel. In: Deutsche Gartenbaugesellschaft/G. Hütten (Hrsg.): Wege zur Naturerziehung, S. 116-133
Probst, W., Hrsg.(1998-2001): Gärten zum Leben und Lernen. Seelze: Kallmeyer (16 Hefte)
Probst, W. (20072): Pflanzen stellen sich vor. Köln: Aulis
Probst, W. (2009): Stoffkreisläufe. Basisartikel in UB 349 (33.Jg), S.2-11. Seelze: Friedrich
Probst, W., Schuchardt,P., Hrsg. (20205): Basiswissen Biologie, Abitur. Berlin: BI-Duden
Purves, W. K. u. a. (201910): Biologie. München: Elsevier
Kadereit, J. W. u.a.(201437): Lehrbuch der Botanik (Ersthrsg. E. Strasburger) Heidelberg: Spektrum
Seyffert, W.: Lehrbuch der Genetik. Spektrum, Heidelberg/Berlin 20032
Taiz, L., Zeiger, L., Møller, I., M., Murphy, A. (2018): Plant Physiology and Development. Oxford: Sinauer
Urry, L. A. u.a. (201911): Campbell Biologie. München:. Pearson Deutschland
Whittaker, R. (1969): New concepts of kingdoms or organisms. Evolutionary relations are better represented by new classifications than by the traditional two kingdoms.. Science 163: pp. 150–160 doi:10.1126/science.163.3863.150. PMID 5762760.














{kind=link}