Huflattich (Tussilago farfara): Wachstum nach der Blüte
(alle Fotos des Beitrags von W. Probst)
Huflattich-Blütenstände, Oberteuringen, 11.3.2017
Die ersten Blütenstände des Huflattichs öffnen sich oft schon im Februar oder Anfang März. Sie sind recht kurzgestielt. Die Stiele verlängern sich etwas im Laufe der Blüte und noch stärker, wenn die Blüten verblüht sind und sich die Köpfchen schließen. Die Achsen biegen sich dann nach unten, sodass die Köpfchen wie kleine Glocken nach unten hängen. Mit der Fruchtreife strecken sich die Blütenstandsachsen wieder und wachsen noch einmal erheblich in die Länge, sodass die Fruchtstände mit den Fallschirmfrüchten sich nun auch über die umgebenden Vegetation erheben und der Wind die Flugfrüchte gut verbreiten kann. Die sehr dünnen, seidigen Haare der Flugschirme haften in nassem Zustand auch sehr gut an vorbeistreifenden Tieren und Menschen. Im April kann man alle Stadien dieser Blütenstandsentwicklung nebeneinander beobachten.
Postflorales Wachstum der Huflattich-Blüten- und Fruchtstände, Oberteuringen, 6.4.2024Blütenstand des Huflattichs mit männlichen und weiblichen BlütenFruchtstand des Huflattichs mit Flugfrüchten
Der Huflattich ist in ganz Eurasien und als invasive Pflanze auch in Nordamerika weit verbreitet. Er ist eine Pionierpflanze auf wechselfeuchten lehmigen Böden und wegen seiner langen, oft über 1 m tief reichenden Ausläufern kommt er zu auch sehr gut mit Hangrutschungen zurecht. Er ist ein sehr typischer Erstbesiedler von frisch aufgeschütteten Straßenböschungen, Hängen von Steinbrüchen und Kiesgruben und anderendurch menschliche Aktivitäten entstandenen Offenböden. An Steilufern von Bächen, Flüssen und Seen, die durch Erosion offengehalten werden, können sich vom Huflattich dominierte Pflanzenbestände lange halten. Ein gutes Beispiel ist die Steilküste der Ostsee an der Flensburger Förde (Abbildung). Die Pflanzensoziologie unterscheidet eine eigene Pflanzengesellschaft, das Poo–Tussolaginetum.
Hangrutschung an der Flensburger Förde mit Huflattich Beständen (Langballigau, 12.3.1992– oben – und 6.5.1990– unten)
Während der deutsche Name auf die hufförmigen Blätter Bezug nimmt, weist der lateinische Name „Tussilago“ auf die hustenlösenden Eigenschaften seiner Inhaltsstoffe hin (lat. tussis = Husten). Deshalb sind Blüten und Blätter auch Bestandteile von Hustentees. Allerdings enthält Huflattich auch karzinogene Pyrrolizidin-Alkaloide , weshalb von der Verwendung selbst gesammelte Huflattichblätter und -blüten abgeraten wird. Mittlerweile gibt es PA-freie Huflattichsorten für die medizinische Verwendung.
Hungerblümchen (Draba verna, syn. Erophila verna)
Die Pflasterfugen in unserem Gartenweg, die im Winter vor allem vom Purpur-Hornzahnmoos begrünt sind, zeigen jetzt im Frühjahr einen ganz besonderen Blütenflor. Tausende winziger Hungerblümchen machen sie zu richtigen Blumenrabatten. Sehr schnell bilden die Zwerge Aus der Familie der Kreuzblütler Früchte und Samen und nach wenigen Wochen sind sie verschwunden. Aber ihre Samen bleiben in den Pflasterfugen und im nächsten Frühjahr, oft schon Ende Februar, treiben sie neue Blütenstände.
Hungerblümchen in Pflasterfugen sind kaum zu sehen, aber wenn man in die Kniee geht, kann man erkennen,dass es sich um richtige kleine Blütenpflanzen handelt.Fotos und Scan W. Probst
In Städten und Siedlungen gibt es viele Ritzen und Fugen. Oft kann man beobachten, wie sie von Anwohnern mühsam ausgekratzt werden. Teilweise kommt es auch zum Einsatz von Herbiziden. Hier könnte man in vielen Fällen Begrünung zulassen und dadurch nicht nur die Biodiversität sondern auch – wie Botaniker der Universität von Santiago de Compostela auch wissenschaftlich bewiesen haben – das Mikroklima verbessern.
An unserem Gartenteich wächst schon seit einigen Jahren eine Schwarz-Erle, die wir regelmäßig sehr stark zurückschneiden, sodass sie nicht höher wird als die krautige Vegetation.
Schwarz-Erle an unserem Gartenteich (11.8.2023, Foto W.Probst)
Heute Morgen (11.9.2023) entdeckte ich an den Blättern eines Zweiges sehr viele Raupen. Die oberen Blätter hatten sie schon ganz kahl gefressen. Sie fraßen die Blätter vor allem vom Rand aus an und bei Störung nahmen sie plötzlich eine auffällige Schrägstellung ein: Sie krümmten ihren Körper bis auf das Hinterende s-förmig nach hinten.
Mithilfe des Internets ließen sich die Raupen eindeutig der Breitfüßigen Erlenblattwespe (Nematus septentrionalis, syn. Craesus septendrionalis) zuordnen, die in ganz Europa vorkommt, aber vor allem in Mittel- und Nordeuropa weit verbreitet ist.
Fressende Raupen der Breitfüßigen Erlenblattwespe (Nematus septentrionalis, syn.Craesus septendrionalis) (16.9.2023, Foto W. Probst) Raupen der Breitfüßigen Erlenblattwespe (Nematus septentrionalis, syn.Craesus septendrionalis) in Schreckstellung (16.9.2023, Foto W. Probst)
Die Weibchen der Blattwespe legen ihre Eier in die Mittelrippen von Blättern, vor allem von Birken, Schwarz-Erlen, Eschen, Ahorn, Weiden und Pappeln. Die die Larven fressen gemeinsam am Blattrand und zwar systematisch bis das Blatt bis zur Mittelrippe und wenigen Seitenrippen abgefressen ist. Laut Wikipedia sollen meist drei Generationen vorkommen, die erste fliegt von Mai bis Juni, die zweite von Juli bis September. Die Puppen der dritten Generation überwintern.
Ein sicheres Unterscheidungsmerkmal zwischen Schmetterlingsraupen und Blattwespenraupen ist die Anzahl der Beinpaare. Schmetterlingsraupen haben maximal acht Beinpaare. Nach den Brustsegmenten folgen bei Schmetterlingsraupen mindestens zwei Segmente ohne Beine. Bei den Raupen der Blattwespen folgt hinter den drei Beinpaaren der Brustsegmente nur ein beinfreies Segment. Alle übrigen Segmente tragen Beinpaare.
Nektarraub
Schon seit Jahrzehnten gedeiht bei mir die afrikanische Aloe aristata. Ich überwintere sie im Zimmer, in der frostfreien Zeit stehen die Pflanzen im Garten.
Aloe aristata im August (Foto W.Probst)
Ich habe auch schon versucht, sie im Freien zu überwintern und einige Male ist es geglückt. Den Freilandaufenthalt danken die Pflanzen mit besonders reichlichen Blüten. Die traubigen Blütenstände können bis zu einem halben Meter lang werden. Oft bilden sie ein bis drei Seitentrauben.
In diesem Topf hat die Aloe an geschützter Stelle zwei Winter überstanden und in den folgenden Sommern besonders reichlich geblüht (Foto W. Probst)
Die mehrere Zentimeter langen, schmalen, orangeroten Blütenkronen sind verwachsen und für heimische Insekten ist der Nektar an der Blütenröhrenbasis kaum zugänglich. In ihrer Heimat werden sie vermutlich von langrüsseligen Nachtfaltern oder sogar von Nektarvögeln bestäubt. In unserem Garten kommt es nur sehr selten vor, dass sich aus einer Blüte eine Frucht entwickelt. Aber der Nektar der Blüten wandert trotzdem in einen Insektenmagen: Wespen, vor allem Feldwespen, knabbern regelmäßig an der Blütenbasis Löcher in die Kronröhre und saugen den Nektar aus.
Eine Feldwespe – vermutlich Polistes dominula – beißt ein Loch in die Kronröhre, um an den Nektar zu gelangen (Foto W. Probst, 9.8.2020)
Aus dem Loch tritt Nektar aus (Foto W. Probst, 16.9.2023)
Nektarraub kann man auch an einheimischen Blütenpfflanzen beobachten, besonders häufig am Akelei, bei dem der Nektar in den Spornern der Blütenblätter gespeichert wird.
Akeleiblüte mit angeknabherten Blütenspornen (Foto W. Pro
Der Stachel-Lattich (Lactuca serriola) trägt wechselständige, stachelige Blätter, die mit ihrer Spreite meist senkrecht stehen und häufig nach Norden bzw. Süden zeigen. Deshalb wird die Pflanze auch „Kompass-Lattich“ genannt. Diese Blattstellung wird als Strahlenschutz gedeutet, da die Pflanze häufig an sehr sonnigen Standorten zu finden ist. Ihr Verbreitungsgebiet reicht von Nordafrika bis in die gemäßigten Zonen.
Die kleinen Fallschirmfrüchte sorgen für eine effektive Windverbreitung. Es handelt sich um eine ausgesprochene Pionierpflanze, die häufig an sonnigen Wegrändern und in lückigen Unkrautfluren zu finden ist.
Der Stachel- oder Kompass-Lattich, eine typische Pionier- und Ruderalpflanze. Foto aus unserem Garten im ersten Sommer nach dem Einzug, 10.7.2008 (Foto W. Probst)
In unserem Garten war sie zu Anfang sehr häufig, vor allem auf einem großen Erdhaufen vom Bauaushub. Später verschwand sie fast vollständig. Zur Zeit keimen ihre Samen vor allem zwischen den Kieseln, die den Plattenweg von der Hauswand trennen.
Nach Ellenberg handelt es sich um eine Volllichtpflanze (L 9) mit geringem Feuchtigkeitsanspruch (F 4) und mäßigem Stickstoffbedarf (N 4)
Junger Kompass-Lattich im Kies zweischen Hausmauer und Terrassenplatten, 18.5.2022 (Foto W. Probst)
Der Grüne Salat (Lactuca sativa) mit den Züchtungen „Kopfsalat“ und „Römischer Salat“ stammt vermutlich vom Stachel-Lattich ab.
Wilde Möhre (Daucus carota) – Fam. Apiaceae
Wilde Möhre (Daucus carota) aus unserem Garten (10.7.2021, Foto W. Probst)
Nach dem Kompass-Lattich möchte ich noch eine einheimische Wildpflanze vorstellen, die Ursprungsart für eine wichtige Kulturpflanze ist: die Wilde Möhre (Daucus carota). Die Pflanze blüht bei uns von Mai bis Anfang August. Dieses Jahr mit seinen vielen warmen Sonnentagen ließ die Möhrenbestände sehr gut gedeihen. Die charakteristischen weißen Doppeldolden, meist mit einer schwarzen „Mohrenblüte“ in der Mitte, bilden an vielen Wegrändern – wie im Bild am Bodensee bei Manzell – große Bestände.
Bestand der Wilden Möhre am Bodenseeufer bei Manzell, 18. Juli 2018(Foto W. Probst)
Dadurch, dass in den letzten Jahren die Wegränder und Straßenränder weniger häufig gemäht werden, haben sich die Bestände dieser Pflanze erheblich vermehrt.
Blütenstand der Wilden Möhre mit „Mohrenblüte“ (29.7.2022, Foto W. Probst)
Innerhalb der Doldenblütengewächse sind weiße Doppeldolden als Blütenstände weit verbreitet. Sie sehen bei den verschiedenen Arten sehr ähnlich aus, aber die Mohrenblüte ist ein Alleinstellungsmerkmal der Wilden Möhre. Über die biologische Bedeutung dieser durch Anthocyane sehr dunkel gefärbten Blüte findet man in der Literatur die Vermutung, dass es sich dabei um eine Fliegenattrappe handelt, die andere Fliegen anlockt.
Man könnte vermuten, dass der Name „Möhre“ oder „Mohrrübe“ mit der dunklen Mittelblüte zu tun hat. Aber das stimmt nicht . Der Name kommt vom mittelhochdeutschen „morche“, „morhe“ oder „more“ für „Rübe“ oder „dicke Pfahlwurzel“. Bis heute werden Möhren in Norddeutschland ja auch „Wurzeln“ genannt. Andere Namen sind Karotte oder Gelbe Rübe. Sie beziehen sich auf die von Carotin verurachte orangegelbe Farbe der Kultur-Möhre. Die Wurzeln der Wilden Möhre sind weißlich oder schwach crremefarben, aber sie schmecken und riechen nach Karotte.
Möhren sind zweijährig. Im ersten Jahr bildet sich eine Blattrosette mit einer dicken Pfahlwurzel. Kultur-Möhren werden in diesem Zustand geerntet. Im zweiten Jahr werden die in der Pfahlwurzel gespeicherten Reservestoffe zum Aufbau der Blütenstände genutzt, die bis über 1 m hoch werden können. Im Gegensatz zu vielen anderen Doldenblütlern sind die Dolden der Möhre vor dem Aufblühen kugelig geschlossen und nach dem Verblühen neigen sich die Doldenäste ebenfalls wieder zu einer Kugel zusammen (an ein Vogelnest erinnernd). Auf den Früchten entwickeln sich hakenförmige Härchen, die der Tierverbreitung dienen.
Kugeliger Fruchtstand der Wilden Möhre, der an ein Vogelnest erinnert (Foto W. Probst)
Blütenstand des Gewöhnlichen Wasserdost (Eupatorium cannabinum) mit Distelfalter (Cynthia cardui), August 1995,(Foto W. Probst)
Der Wasserdost ist eine ausgesprochene Schmetterlingspflanze. In unserem Garten in Flensburg wurde die Staude regelmäßig von vielen Faltern besucht. In unserem Garten in Oberteuringen hat sich der Wasserdost von selbst am Teichufer eingefunden. Die leichten Flugfrüchte werden weit verbreitet. Allerdings ist die Schmetterlingsfauna in den letzten zwei Jahrzehnten sehr viel ärmer geworden, meistens konnten wir deshalb nur einzelne Falterbesuche beobachten.
Gartenteich mit fruchtenden Wasserdost-Pflanzen im September (Foto W. Probst)
Wasserdost-Arten stammen aus Afrika, Asien, Nord- und Südamerika, nur wenige sind in Europa heimisch. Für die Gartenkultur werden aber mehrere Arten angeboten, zum Beispiel die Hochstaude Eupatorium fistulosum (Purpur-Wasserdost, Röhriger Wasserdost) aus Nordamerika, von dem es eine Reihe von Zuchtformen gibt.
„Dost“ bedeutet im Mittelhochdeutschen „etwas buschiges, etwas, das in Büschen wächst“. Ohne Zusatz „Wasser“ bezeichnet Dost die Pflanzengattung Origanum aus der Familie der Lippenblütler. Dazu gehören Majoran (Origanum majoranum) und Echter oder Gewöhnlicher Dost, italienisch Oregano (Origanum vulgare), der mit dem Wasserdost nur die violette Blütenfarbe gemeinsam hat. Dies gilt auch für den Lippenblütler Wirbeldost (Clinopodium vulgare).
Die Blätter des Gewöhnlichen Wasserdosts sind drei geteilt und erinnern entfernt an Hanfblätter (Cannabis), daher das Art-Epitheton „cannabinum“.
Seerosen-Honig
Auf den Seerosenblättern in meinem Teich kann man ziemlich regelmäßig jedes Jahr zwei Insektenarten beobachten.
Seerosenblätter mit Spuren des Seerosen-Blattkäfers (Galerucella nymphaeae) (Foto W. Probst)
Zum einen ist das der Seerosen-Blattkäfer (Galerucella nymphaeae). Alle Lebensstadien, Eier, Larven, Puppen und Imagines des Käfers leben auf den Blättern von Seerosen. Außerdem kommen die Käfer auf Gelber Teichrose, Wasser-Knöterich und Pfeilkraut, eigenartigerweise auch auf Erdbeerpflanzen vor. Die unregelmäßigen Rillen, die Larven und Käfer in die obere Blattschicht fressen, können bei Massenbefall sehr auffällig sein und die Blätter auch zu frühem Absterben bringen, untergehen tun sie allerdings nicht so schnell, da die untere Epidermis nicht angefressen wird.
Seerosenblatt mit Seerosen-Blattlaus (Rhopalosiphum nymphaea) und Honigbiene (Foto W. Probst)
Die zweite Insektenart, die auf den Seerosenblättern zum Teil massenhaft auftreten kann, ist die Seerosen-Blattlaus (Rhopalosiphum nymphaea). Ihr Honigtau kann die Spaltöffnungen auf der Seerosenblattoberseite verstopfen und deshalb die Blätter zum vorzeitigen Absterben bringen. Außerdem lockt der süße Saft auch Insekten, Fliegen und vor allem Bienen, an. Sie verwandeln die süßen Ausscheidungen in Honig, Seerosen-Honig.
Im Mai 2023 hat sich eine großeStaude des Färber-Waid (Isatis tinctoria) zwischen Mülleimerplatz und Komo´post entwickelt (Foto W. Probst)
Auch 2023 gibt es wieder einen Spontanzugang an unserem Kompost- und Mülleimer-Stellplatz: Es hat sich eine prächtige fast 2 m hohe Staude des Färber-Waids (Isatis tinctoria) entwickelt. Mir fiel schon letztes Jahr die große, grundständige Blattrosette auf, aber ich konnte sie nicht zuordnen.
Diese schon seit vorgeschichtlichen Zeiten genutzte Kulturpflanze aus der Familie der Kreuzblütler (Brassicaceae) war bis ins 16. Jahrhundert wichtiger Lieferant für den blauen Farbstoff Indigo, der vor allem für das Färben von Leinenstoffen aber auch für Holzanstriche verwendet wurde. Erst nachdem der Echte Indigo (Indigofera tinctoria) aus Indien, China und Afrika eingeführt wurde, verlor Färber-Waid schnell an Bedeutung.
Nun findet man die Pflanze in Mitteleuropa immer noch, vor allem an kalkreichen, relativ trockenen Standorten. Aber wie er in unseren Garten gekommen ist, bleibt eine offene Frage, denn in der näheren Nachbarschaft kenne ich keine Standorte.
Färber-Waid (Isatis tinctoria),Maintal, Juni 1987 (Foto W. Probst)
Allerdings werden Färber-Waid-Samen auch vom Gartenversandhandel angeboten. Die Pflanze gilt als gutes Bienenfutter: Die Samen entwickeln sich in einsamigen „Schötchen“, die sich aber – im Gegensatz zu den für die übrige Kreuzblütler typischen Schoten und Schötchen – nicht öffnen, botanisch gesehen also eigentlich Nüsse sind.
Der Himmel über Oberteuringen
5.November 2022 (Foto W. Probst)
Am 5. November 2022, während in Sharm-EL-Sheikh die 27. UN-Klimakonferenz tagte, hatten wir in Oberteuringen einen wunderschönen Sonnentag mit strahlendblauem Himmel. So sah dieser Himmel aus, als ich morgens um neun auf unserer Terrasse rollte: Alle erkennbaren Wolkenbildungen auf diesem Foto gehen auf Kondensstreifen von Flugzeugen zurück. Für jedes Wassermoleküle, dass diese Kondensstreifen verursacht, wird auch ein Kohlenstoffdioxidmolekül ausgestoßen. Die Treibhausgasemissionen des Luftverkehrs werden dadurch indirekt sichtbar.
„Das Unheil im Kondensstreifen bekommt keine Gestalt, obwohl jedes vernünftige westeuropäische Bewusstsein von diesem Unheil weiß. Kondensstreifen gibt es nur, weil es den Flugverkehr gibt, und der Flugverkehr ist ein sichtbares Zeichen für die globale Erwärmung. Ein Kondensstreifen ist eine Beschädigung der Atmosphäre durch den Menschen. Wenn wir diese Beschädigung denken und ihre Folgen bedenken, müsste er einen anderen Namen bekommen. Kondensstreifen sind anthropogene Wolken, was als Begriff wiederum in die Vergangenheit weist. In die Zukunft, in die Folgen gedacht, müssten sie einen Namen des Unglücks bekommen, ein Name der Schmerzen und der Schönheit. Es bleibt eine Wolke, weiß und hell und glänzend am Himmel, ein freundliches Zeichen des Untergangs im Abendlicht.“
Bärfuss, Lukas. Vaters Kiste: Eine Geschichte über das Erben (S.62-63). Rowohlt E-Book. Kindle-Version.
Schildblatt (Darmera peltata) im Frühjahr, 2.5.2013 (Foto W. Probst)
Das Schildblatt (Darmera peltata) stammt aus dem Westen der Vereinigten Staaten von Amerika. Es gedeiht an Bach- und Flussufern in den Staaten Kalifornien und Oregon. Nach unserem Einzug in Oberteuringen erhielten wir einige Pflanzen von unserer Nachbarin. Sie wuchsen zunächst an einigen Stellen in unserem Garten aber dauerhaft hielten sie sich nur am Rand unseres Gartenteiches.
Natürlich, es handelt sich um einen Exoten und strenge Verfechter der Naturgartenidee plädieren dafür, nur Einheimische im Garten wachsen zu lassen. Ich gehöre nicht dazu. Exotische Pflanzen, die mich faszinieren und die im Garten gut gedeihen dürfen dort auch gerne wachsen. In unserem Garten gehört das Schilfblatt dazu.
Schon im April, vor den Blättern, erscheinen die kugeligen rosa Blütenstände an einem Stiel, der bis 30 cm hoch werden kann. Erst im Mai entfalten sich dann die großen Schildblätter. Die Blütenstände strecken sich bis zur Fruchtreife noch einmal auf doppelte Länge, insbesondere die Achsen zwischen den einzelnen Blüten verlängern sich und in jeder Blüte entwickeln sich 2-3 der für Steinbrechergewächse typischen Bälgchen.
Blick in den Blütenstand des Schildblatts, 2.5.2013 (Foto W. Probst)Blütenstand des Schildblatts auf dem Weg zur Fruchtreife, 9.5.2019 (Foto W. Probst)Sich entfaltendes Blatt, Mai 2019 (Foto W. Probst)
Vom Lebensformentyp ähnelt das Schildblatt unserer Gewöhnlichen Pestwurz (Petasites hybridus), die ebenfalls bevorzugt an Bach- und Flussufern und auf teilweise überfluteten Kiesbänken gedeiht und die im zeitigen Frühjahr ihre violetten Blütenstände treibt, bevor sich die riesigen rhabarberartigen Blätter entwickeln. Allerdings würde sich die Pestwurz an unserem nur mit Regenwasser gespeisten und deshalb sehr nährmineralarmen Teich vermutlich nicht gut entwickeln. Auch das Schildblatt erreicht hier keine maximalen Größen – ich sehe das im Vergleich zu den Exemplaren bei unserer Nachbarin, die einen sehr eutrophen Teich mit sehr vielen Fischen pflegt. Trotzdem spenden auch bei uns die Schildblätter den Sommer über sehr viel Schatten. Darunter gedeihen fast nur noch Moose.
Im Herbst erhöht das Schildblatt noch einmal seine Attraktivität durch eine ausgezeichnete Herbstfärbung. Im Winter zieht sich die Pflanze ganz in das kräftige unterirdische Rhizom zurück.
Herbstfärbung des Schildblatts, 25.10.2023 (Foto W. Probst)
Minen, die nicht explodieren
Akeleipflanze mit vielen Minen (Foto W.Probst 26.10.2023)
Ich beobachte eigenartige Linien und Muster, mäandererartig verschlungene Pfade, auf Akeleiblättern. Wenn ich die Blätter gegen das Licht halte, wird deutlich, dass die Blätter an diesen Pfaden sehr durchscheinend sind. Sie bestehen nur aus den chlorophyllfreien Epidermen, den einzellschichtigen Häutchen, die das Blatt nach außen abschließen. Das Blattinnere, das grüne Mesophyll, fehlt. Es wurde von kleinen Insektenlarven aufgefressen, die sich durch das Blattinnere bohren.
Man nennt diese durch Fraß entstandenen Gänge „Minen“ oder „Hyponomien“. Es gibt eine sehr große Anzahl unterschiedlicher Blattminen, die von Mücken-, Fliegen-, Kleinschmetterlings-, Käfer- und Blattwespenlaven erzeugt werden können.
Scan eines Akleiblattes mit Gangminen von Phytomyza aquilegivora (30.10.2023, W. Probst,)
Bei den abgebildeten Akeleiblättern ist der Verursacher die Akelei-Minierfliege Phytomyza aquilegivora. Den Minentyp, bei dem sich die Insektenlarven einen Gang entlang fressen, dessen Durchmesser mit dem Wachstum der Larve immer breiter wird, nennt man „Gangminen“. Eine nahe verwandte Minierfliegen-Art, Phytomyza aquilegiae, frisst sich flächig durch das Blatt und bildet „Platzminen“.
Platzminen voon Phytomyza aqulegiae (7.10.2015, Foto W. Probst)
Als Pionier der Pflanzenminen-Forschung gilt Erich Martin Hering und sein Werk „Die Blattminen Mittel- und Nordeuropas einschließlich Englands“. Die Bestimmung ist mit dieser umfassenden Monographie sehr gut möglich, da die Minen nach ihren Wirtspflanzen sortiert sind.
Die Minen an Akeleiblättern sind verhältnismäßig häufig, andere Pflanzen, die oft von Minierern befallen werden, sind zum Beispiel Geißblatt bzw. Heckenkirsche, Springkraut, Gänsedistel und Klette.
Gangminen von Larven der Geißblatt-Minierfliege (Phytomyza xylostei) auf dem Blatt der Roten Heckenkirsche (Lonicera xylosteum) (8.1996, Foto W. Probst)
Orchideen am Gartenteich
An unserem Gartenteich haben sich „freiwillig“ zwei Orchideenarten eingefunden. 2016 konnte ich am Teichufer zum ersten Mal eine blühende Fleischfarbene Fingerwurz (Dactylorhiza incarnata) entdecken. Die Art kommt in einem etwa 1 km entfernten Naturschutzgebiet (Altweiherwiese) vor. In den Folgejahren hat sich die Anzahl der Orchideenpflanzen langsam aber kontinuierlich erhöht.
Dactylorhiza incarnata Mit der Kleinlibelle Coenagrion puella (20.5.16; Foto W. Probst)
Zu Dactylorhiza incarnata kam noch die Breitblättrige Fingerwurz (Dactyloriza majalis), die ebenfalls im NSG Altweiherwiese zu finden ist. 2022 konnte ich 14 PflanzenDieser beiden Arten zählen, dieses Jahr (2023) ging die Anzahl auf 11 zurück. Dabei hat eventuell Schneckenfraß eine Rolle gespielt.
Dactylorhiza majalis (1.5.2020, Foto W. Probst)
Unser Teich wird immer wieder mit Regenwasser aus einer Zisterne aufgefüllt und ist sehr mineralstoffarm. Die Pflanzen am Teichufer – zum Beispiel Blutweiderich und Wasserdost – bleiben ziemlich klein. Dazwischen gedeihen Moose sehr gut, insbesondere das Spießmoos (Calliergonella cuspidata). Aus diesen Moosrasen sprießen die Orchideen.
Binsen-Schmuckzikade (Cicadella viridis)
Binsen-Schmuckzikade (Cicadella viridis) -(14.10.22; Foto W. Probst)
Im Oktober 2022 beobachtete ich an den schon fast abgestorbenen Stängeln einer Wald-Engelwurz (Angelica silvestris) am Rand meines Gartenteiches etwa 0.7cm lange, hell türkisgrünliche Zikaden. Mithilfe des Internets konnte ich sie als Binsen-Schmuckzikade (Cicadella viridis) identifizieren. Bei den von mir beobachteten Tieren handelte es sich um Weibchen, die Männchen haben laut Wikipedia blaue Flügeldecken. Charakteristisch sind zwei ziemlich große schwarze Flecken zwischen den Komplexaugen.
Wie der Name schon sagt, soll die kleine Zikate vorwiegend an Binsen saugen, aber sie nimmt auch den Xylemsaft von vielen anderen Pflanzenarten auf. Um genügend Nährstoffe zu erhalten, müssen die kleinen Zikaden ziemlich große Mengen des sehr wässrigen sie Xylemsaftes einsaugen, die überflüssige Flüssigkeit wird dann – wie im Foto zu sehen – in großen Tropfen abgeschieden.
In Mitteleuropa sollen die Binsen-Schmuckzikaden pro Jahr ein bis zwei in Südeuropa drei und mehr Generationen bilden. Imagines der hemimetabolen Zikaden kann man bei uns von Mai bis Oktober beobachten.
Schmuckzikaden (Cicadellinae) sind eine Unterfamilie der Zwergzikaden (Familie Cicadellidae). Die wegen ihrem auffällig schaumigen Larvenschutz („Kuckucksspeichel“) bekannten Schaumzikaden (Familie Cercopidae) sind eine Schwestergruppe der Schmuckzikaden.
Vom Menschen unberührte Natur macht derzeit weniger als ein Viertel der Erdoberfläche aus. Den Forderungen, solche Flächen zur Stabilisierung des Bioplaneten zu vergrößern, steht die wachsende Weltbevölkerung und die auf Wachstum begründete Weltwirtschaft entgegen. Gibt es trotzdem Möglichkeiten, natürliche Funktionsabläufe zu vermehren?
Wildnis und Naturschutz
Die vom Menschen noch kaum veränderten Gebiet der Erdoberfläche machen gegenwärtig weniger als ein Viertel aus. 77% der Landfläche (ohne Antarktika) und 87% der Meere sind bis heute durch menschliche Aktivitäten verändert worden, der größte Teil davon in den letzten 50 Jahren (Watson, Allen u.a. 2018). Dies wird von vielen Ökologen als ein großes Problem angesehen, denn vom Menschen bisher kaum beeinflussten Wildnis-Gebiete gelten als wichtigster Puffer gegen den Verlust der biologischen Vielfalt und die Klimaveränderungen. Wildnisgebiete regulieren Wasserkreisläufe und Klimazyklen und schützen damit vor extremen Wetterereignissen. Außerdem stellen sie wichtige Referenzflächen für die Regeneration und Renaturierung degradierter Landflächen und Meeresgebiete dar. Die Degradation und Fragmentaktion naturnaher Restflächen verstärken die nachteiligen Auswirkungen der Klimaerwärmung auf die Biodiversität (Mantyka-Pringle u. a. 2012).
Den Erhalt von Wildnis ist deshalb ein wichtiges Naturschutzziel.
Aber was ist Wildnis? Ist es im Sinne Aldo Leopolds von Menschen unberührte Natur? Oder sind mit domestizierten Rindern und Pferden beweidete „halboffene Weidelandschaften“ ebenso Wildnis, wie dies Jan Haft in seinem Buch „Wildnis“ darstellt? Welche Rolle spielt Wildnis für die Biodiversität, für den Klimaschutz und für den Erhalt natürlicher Ressourcen? Haben Aufforstungsprogramme etwas mit Wildnis zu tun? Inwiefern ist der Naturschutz mit Wildnis-Vorstellungen verknüpft?
Viele Fragen. Ein Versuch, sie zu beantworten, lässt schnell erkennen, dass es recht unterschiedliche menschliche Vorstellungen von „wilder Natur“ und den Beziehungen der Menschen zu solcher Wildnis gibt.
Europäische Wildnis?
Die in Mitteleuropa seit der letzten Kaltzeit in etwa 12 000 Jahren – also einer erdgeschichtlich sehr kurzen Zeitspanne – entstandenen Landschaften waren von Anfang an vom Menschen beeinflusst. Die menschliche Nutzung hat ein kleinräumiges Mosaik von Lebensräumen geschaffen und zu einer Artenvielfalt geführt, die sich vermutlich ohne den Menschen und seine Nutztiere nicht oder zumindest nicht so schnell entwickelt hätte.
Eine kleinräumig strukturierte Kulturlandschaft hat sich in Mitteleuropa bis heute in einigen Gebieten erhalten (Foto W. Probst 14.9.2012)
Ein flächendeckender Urwald, wie er über die Jahrhunderte heute vermutlich ohne menschlichen Einfluss in Mitteleuropa entstehen würde, hätte sicher eine geringere Artenvielfalt aufzuweisen als die ursprüngliche, vorindustrielle Kulturlandschaft. Der Biologe und Naturfilmer Jan Haft belegt dies in seinem Buch „Wildnis“ mit gut recherchierten Zahlen und Aussagen von Experten (Haft 2023). Es ist deshalb verständlich, dass Naturschutz in Mitteleuropa in vielen Fällen mit Managementmaßnahmen verbunden ist, bei denen es darum geht, traditionelle Landbewirtschaftungsmaßnahmen nachzuahmen. Schilfbestände in Feuchtgebieten werden abgemäht und das Mähgut gut wird entfernt um einen Zustand magerer Feuchtwiesen zu erreichen, der alten Streuwiesen entspricht. Heiden und Moore werden maschinell oder von Hand von Gehölzen befreit (entkusselt), um einen Zustand herzustellen, der einer extensiven Beweidung entspricht. Feldhecken, die früher auch der Nutzholzgewinnung dienten, werden als Naturschutzmaßnahme weiterhin regelmäßig „auf den Stock gesetzt“, um das Durchwachsen zu Baumreihen zu verhindern und den für Kleinsäuger, Vögel, Reptilien und viele Wirbellosen wertvollen Heckencharakter zu erhalten. Alle diese Maßnahmen zielen auf den Erhalt von Landschaften ab, die man nicht als „unberührte Natur“ bezeichnen kann.
In den zwischeneiszeitlichen Warmzeiten allerdings war die Biodiversität ebenfalls deutlich höher. Ursache waren vermutlich die zahlreichen großen Herbivoren, deren Weidetätigkeit die Bildung geschlossener Urwälder verhinderte. Vielmehr herrschten offene, savannenähnliche Landschaften , wie sie heute zum Beispiel noch in Afrika zu finden sind. Dass es solche großen Pflanzenfresser seit dem Ende der letzten Kaltzeit in Europa nicht mehr gibt, ist vermutlich auf die Tätigkeit menschlicher Jäger zurückzuführen ( Sandom et al. 2014). Streng genommen könnte man deshalb diese voreiszeitliche Landschaft als die eigentliche mitteleuropäische Wildnis ansehen.
Nordamerikanische Wilderness
In Nordamerika ist der Naturschutz deutlich stärker mit dem Wildnisbegriff im Sinne von unberührter Natur verbunden als in Europa. Der Naturalist und Dichter Henry David Thoureau forderte schon 1862, dass jede amerikanische Stadt zur Bildung und Erholung ihrer Bevölkerung 200-400 ha Wildnis so bewahren sollte, dass darin nicht einmal die Spur eines geschnittenen Stockes zu erkennen wäre (nach Trommer 2023). Auch für den großen amerikanischen Naturschützer John Muir war die wilde, von Menschen unberührte Natur der zu schützende Idealzustand. Ebenso setzte sich der Wildtierbiologe Aldo Leopold (1887-1948) für die Bewahrung von Wildnis als einem von Menschen weitestgehend unbeeinflusstem Naturraum ein. Seine Schriften hatten großen Einfluss auf den 1964 beschlossenen Wilderness Act, mit dem ein System von vollständig geschützten Wilderness Areas geschaffen wurde (Henderson o.J.).
Diese unterschiedlichen Vorstellungen von Naturschutz in Nordamerika und Europa hängen sicherlich auch damit zusammen, dass die Landschaftsveränderungen in Nordamerika im 18. und vor allem im 19. Jahrhundert in atemberaubender Geschwindigkeit verliefen und deshalb im Laufe eines Menschenlebens sehr gut zu beobachten waren. Die europäischen Siedler bewirkten eine sehr rasche und drastische Veränderung und verhinderten von vorneherein die Entwicklung einer europäischen Verhältnissen vergleichbaren kleinräumig strukturierten Kulturlandschaft.
Agrarlandschaft in Illinois (Foto W.Probst 1989)
Außerdem war der Ausgangszustand nach der Eiszeit in Nordamerika biodiverser als in Europa. In Nordamerika konnten sich die Biodiversität nach der letzten Eiszeit schneller regenerieren als in Europa, da die Biozönosen während der Kaltzeiten wegen der vorwiegend von Norden nach Süden streichenden Gebirge nicht so stark dezimiert wurden. In Mitteleuropas war eine Rückzugsmöglichkeit nach Süden durch die Alpen weitgehend versperrt.
Allerdings sind auch in Nordamerika viele der vor den Kaltzeiten oder in Zwischenwarmzeiten noch existenten großen Pflanzenfesser einschließlich ihrer Prädatoren verschwunden. Es ist naheliegend, zu vermuten, dass auch hier menschlicher Einfluss, die Jagd, für das Aussterben entscheidend war. Ähnliche Entwicklungen kann man auch für Australien und Teile Asiens nachweisen. Lediglich in Afrika haben bis heute eine Vielzahl großer Herbivoren und Carnivoren überlebt. Dies wird damit in Verbindung gebracht, dass sich in Afrika Menschen und Großsäuger über lange Zeiträume parallel entwickelt haben.
Welche Wildnis wollen wir?
Aus diesen Überlegungen wird deutlich, dass nicht so ganz eindeutig ist, was jeweils unter „Wildnis“ , also einem ursprünglichen Naturzustand, gemeint ist und welche günstigen Wirkungen auf eine nachhaltige Entwicklung des Bioplaneten Erde sich daraus ergeben. Geht es um einen Zustand ohne jeglichen menschlichen Einfluss, also um Ökosysteme ohne Homo sapiens oder gehören auch sogenannte Naturvölker dazu? Welche Rolle spielen reich strukturierte Kulturlandschaften, wie sie bis zu Beginn der Industrialisierung in Europa vorherrschend waren? Wie sind die Veränderungen – man kann auch sagen Ausrottungen – zu bewerten, die schon durch Jäger und Sammler bei der Besiedelung Australiens und Amerikas bewirkt wurden? Wo zieht man die Grenzen? Ist es wirklich notwendig, völlig unberührte (menschenfreie) Natur zu erhalten, oder können menschliche Aktivitäten teilweise dazu führen, dass Funktionen im Naturgeschehen wieder ablaufen, die vormenschlichen Bedingungen entsprechen? Geht es also mehr um „wilde“ Funktionsabläufe als den Erhalt eines menschenfreien Zustandes?
Wilde Weiden
Heckrinder-Bulle im Leimbach-Hepbacher Ried bei Markdorf, Baden-Württemberg (Foto Probst 2011)
Jan Haft zielt in seinem Buch „Wildnis“ genau auf dieses Funktionsverständnis von Wildnis ab, das im Naturschutz auch als „Prozessschutz“ bezeichnet wird. Dabei geht es ihm vor allem um die Ökosysteme mit großen Pflanzenfressern, die in vielen Gebieten der Erde vor dem Erscheinen des Menschen große Räume einnahmen. Diese vorzeitliche Wildnis könnte funktional wiederhergestellt werden durch domestiziert Weidetiere, deren Populationen nicht durch Carnivoren sondern durch den Menschen reguliert werden. Die mittlerweile an vielen Orten etablierten „halboffenen Weidelandschaften“ sind ein gutes Beispiel dafür, dass solche wilde Weiden der Biodiversität wirklich sehr förderlich sind und dass in solchen Gebieten viele bedrohte Arten sich wieder ausbreiten und regenerieren konnten. Zwei sehr gut dokumentiertes Beispieleaus meiner früheren Heimat sind die auf einem ehemaligen Truppenübungsplatz der Bundeswehr entstandene Weidelandschaft „Stiftungsland Schäferhaus“ bei Flensburg und das Stiftungsland Winderatter See – Kielstau (Janßen 2011-2020)
Das Prinzip dieser Art von Verwilderung lässt sich auf andere Bereiche ausweiten. Einige Beispiele:
Aufforstung
Bäume pflanzen und durch Trockenheit und Schädlingsbefall – vor allem Windbruch und Borkenkäfer – geschädigte oder zusammengebrochenen Wälder durch Aufforstung zu regenerieren gilt nicht nur als eine wichtige Maßnahme des Klimaschutzes sondern auch des Naturschutzes und der Förderung der Biodiversität. Dem widerspricht zum Beispiel der Förster und Erfolgsautor Peter Wohlleben: „Wald kommt von ganz alleine zurück, das macht er seit 300 Millionen Jahren.“ Global gäbe es kein Beispiel dafür, dass gepflanzter Wald besser funktioniert, als ein Wald, der von selbst zurück wächst. Besonders widerspricht Wohlleben der Annahme, Bäumepflanzen sei eine unumstrittene Klimaschutzmaßnahme. Eine frisch gepflanzte Aufforstung stoße in den ersten Jahren bis Jahrzehnten mehr CO2 aus, als die neu gepflanzten Bäume aufnehmen könnten (Wohlleben in“Hart aber fair“ , 01.11.21).
Erfahrungen im Nationalpark Bayerischer Wald geben Wohllebens Auffassung recht. Nachdem in den 1990 er Jahren durch Borkenkäferbefall rund 60.000 ha Wald zugrunde gegangen waren, hielt die Nationalparkverwaltung trotz großer Proteste der Öffentlichkeit an ihrer Nichteingriffsstrategie fest. Die sich hervorragend regenerierenden Bergwaldflächen sind mittlerweile ein international bekanntes Beispiel für natürliche Waldregeneration (Bibelriether 2017).
Ackerbau
Die hohe Biodiversität einer kleinräumig strukturierten Kulturlandschaft, wie sie in früheren Jahrhunderten für Mitteleuropa typisch war, ist unbestritten. Viele hiesige Naturschutzmaßnahmen zielen deshalb darauf ab, alte bäuerliche Bewirtschaftungsformen zu simulieren. Dies geht aber nur auf verhältnismäßig kleinen, abgeschlossenen Naturschutzflächen. Großflächig dominieren weiterhin große, unstrukturierte Ackerflächen, da nur solche mit Großmaschinen rationell bearbeitet werden können. Wäre es nicht denkbar, dass eine zunehmende Digitalisierung der Landwirtschaft auch eine rationelle maschinelle Bearbeitung kleinräumig strukturierte Anbauflächen ermöglichen würde? Statt dinosaurierartiger Riesenmaschinen könnten kleine Agrarroboter Bearbeitung und Ernte übernehmen, die von Satelliten oder Drohnen gesteuert ganz gezielt eingesetzt werden könnten. Sie würden sich an einem verhältnismäßig engmaschigen Netz von Feldhecken und Feldgehölzen, Randstreifen und Saumbiotopen nicht stören. So könnte eine kostengünstige Produktion ermöglicht werden, ohne natürliche Funktionsabläufe vollkommen zu unterbinden.
Auch die arbeitsintensiven Methoden der Permamakulturen und der Agroforestry, die versuchen, natürliche Prozesse nicht zu unterdrücken sondern auszunutzen, könnten durch KI-Einsatz rentabler werden.
Landwirtschaft, die natürliche Funktionsabläufe zulässt (Grafik W. Probst)
KI in der Landwirtschaft
„Der nächste Schritt in der technologischen Entwicklung intelligenter landwirtschaftlicher Maschinen könnte eine Art Schweizer Armeemesser sein: ein Roboter, der jede Pflanze individuell behandelt, nicht nur mit Herbiziden sondern auch mit angepassten Düngemitteln, Insektiziden und Fungiziden und gezielter Bewässerung, alles in einem Arbeitsgang und jeweils nur in der benötigten Menge. Die Folgen einer solchen. Behandlung von Einzelpflanzen statt von ganzen Feldern bedeutet nicht nur eine deutliche Reduktion benötigter Chemikalien und anderer Ressourcen. Es könnte schließlich auch zu einem Ende der Monokulturen führen, einem Ende von Kornfeldern oder Sojafeldern soweit das Auge reicht, die heute der Normalfall sind. Monokulturen laugen Böden aus und sind riskant, da solche nur von einer Pflanzenart bewachsene Felder für Schädlingsbefall und andere Katastrophen besonders anfällig sind.“ (Übersetzt aus Little, A. (2019): The fate of food. What we’ll eat in a bigger,hotter,smarter World. London: Oneworld Publications, p.106)
Paludikultur
Bis vor 200 Jahren waren Torfmoore die letzten unberührten Naturlandschaften Mitteleuropas. Durch Entwässerung und Bodenbearbeitung, Torfstich zur Brennmaterialgewinnung und später für Blumenerde und Gärtnereibedarf führten zum weitgehenden Verschwinden ursprünglicher Moore mit aktiver Torfbildung. Im Zuge der Klimaerhitzung hat man festgestellt, dass die Torfbildung unter Mooren eine sehr effektive Form der Kohlenstoffspeicherung darstellt. Deshalb werden seit einiger Zeit große Anstrengungen unternommen, um aktive Moore zu regenerieren. Dies muss aber nicht unbedingt zur Herstellung des ursprünglichen Zustandes führen. Eine Alternative sind die sogenannten Paludikulturen, bei denen auf wieder vernässten Torfböden nutzbare Pflanzenproduktion betrieben wird. Geerntet werden können nicht nur Schilf und Sauergräser sondern auch Torfmoose, aus denen ein für Gärtnereizwecke besonders wertvolles, dem Hochmoortorf entsprechendes Grundsubstrat gewonnen werden kann. Die Kohlenstoff-speichernden Torfschichten bleiben erhalten. Auch weitere ökologische Funktionen wie Regulierung des Wasserhaushaltes und Erhalt von Lebensräumen für moortypische Tiere und Pflanzen blieben – zumindest teilweise – erhalten (Tanneberger, Schroeder 2023)
Migration
Arten, die sich in einem Gebiet ausgebreitet und etabliert haben, in dem sie zuvor nicht heimisch waren, nennt man Neobiota (auch Neobionten, Sing.der Neobiont). Enger gefasst versteht man darunter nur solche Arten, für deren Einbürgerung indirekt oder direkt menschliche Aktivitäten verantwortlich waren. Arten, die sich ohne menschlichen Einfluss ausgebreitet haben, werden dann als Neueinheimische (Neonative) bezeichnet. Besonders wichtig für Neobiota im engeren Sinne ist der weltweite Güterverkehr.
Nach einer Recherche von Kleunen et al. 2015 wurden bs dahin weltweit 13.168 Pflanzenarten durch menschliche Aktivitäten in neuen Gebieten eingebürgert. Besonders neobiontenreich ist Nordamerika, die größte Anzahl der weltweit neu eingebürgerten Arten stammt aus Europa. Beides hängt vermutlich direkt mit der Kolonisation zusammen, die von Europa ausging.
Vom Naturschutz wird diese menschenbedingte Migration zumeist als großes Problem angesehen, da neu eingewanderte Arten etablierte, heimische Arten verdrängen und Ökosysteme verändern können. Der Naturschutz versucht deshalb, diese Migration zu verhindern und die Migranten wenn möglich wieder aus den neu eroberten Gebieten zu verdrängen. Tatsächlich haben Neobiota teilweise zu drastischen Veränderungen der ursprünglichen Ökosysteme beigetragen. Dies gilt besonders für pazifische Inseln, die von europäischen Kolonisatoren nicht nur mit landwirtschaftlichen Nutzpflanzen und Nutztieren (Schweine, Ziegen) sondern auch mit Ratten und europäischen Wildpflanzen von Äckern und Weiden „geimpft“ wurden. Die sehr speziellen Ökosysteme hatten solchen im wahrsten Sinne des Wortes invasiven Arten nichts oder wenig entgegenzusetzen und viele auf den Inseln endemisch Arten wurden ausgerottet.
Andererseits ist Migration ein sehr natürlicher Vorgang, der für die Geschichte des Lebens auf der Erde eine entscheidende Rolle gespielt hat. Mancuso (2021) bezeichnet Migration nicht ganz zu Unrecht sogar als „Essenz des Lebens“. Allen Lebewesen, so Mancuso, sei ein „Wandertrieb“ eigen, das Bestreben, sich möglichst effektiv auszubreiten, das Verbreitungsareal zu vergrößern. Durch solche Wanderungen bedingte Veränderungen wären für die Entwicklung des Lebens auf unserem Planeten – nicht zuletzt auch für die Evolution des Menschen – von großer Bedeutung. Vom Menschen geförderte oder verursachte Migration ist nicht etwas grundsätzlich anderes als natürliche Migration, allerdings kann vom Menschen geförderte Ausbreitung natürliche Ausbreitungsschranken schneller überwinden und auch große Entfernungen können durch moderne Verkehrsmittel schnell überbrückt werden.
Um den Artenbestand von Inseln zu erklären, haben MacArthur und Edward O. Wilson 1967 die mittlerweile breit akzeptierte Gleichgewichtstheorie der Inselbesiedelung entwickelt. Danach stellt sich – qualitativ leicht zu beschreiben – auf jeder Insel ein Gleichgewicht zwischen Einwanderungsrate und Aussterberate der Arten ein. Je mehr Arten auf einer Insel vorhanden sind, desto geringer ist die Einwanderungsrate. Entweder, da keine Arten zur Einwanderung mehr zur Verfügung stehen, oder, da es keinen Platz mehr für die neu zugekommenen Arten gibt, da also keine „Nischenbildung“ mehr für sie möglich ist. Umgekehrt ist die Aussterberate umso größer, je mehr Arten auf der Insel sind. Steht genügend Zeit zur Verfügung, stellt sich ein Gleichgewicht ein, eine bestimmte Artenanzahl. Die Zusammensetzung der Arten, das Artenspektrum, kann sich oder muss sich allerdings weiter ändern, da ja immer Arten aussterben und Arten einwandern, jeweils in einer Rate, die dem Gleichgewicht entspricht. Ohne Migration würde die Artenanzahl auf Inseln danach kontinuierlich abnehmen. Dies gilt aber natürlich auch für andere mehr oder weniger abgeschlossene Gebiete und vermutlich sogar für ganze Kontinente.
Die meisten Neobiota haben sich gut in die Ökosysteme integriert, ohne dass nachteilige ökologische Auswirkungen erkennbar wären. Eine gezielte Bekämpfung ist deshalb in den meisten Fällen nicht notwendig und – wenn sich die Arten schon weit verbreitet haben – auch wenig erfolgversprechend. Die Ausbreitung und Etablierung von Neobiota kann bei sich veränderndem Klima sogar eine Stabilisierung von Ökosystemen bedeuten. Auch das Bundesamt für Naturschutz empfiehlt deshalb eine weitgehende Akzeptanz der Neubürger und eine Bekämpfung nur in begründeten Einzelfällen.
Verkehr
Die Hauptprobleme, die sich durch privaten und öffentlichen Verkehr ergeben, sind die Zerschneidung der Landschaft und die Produktion schädlicher Abgase. Das zweitgenannte Problem versucht man durch „grüne Energie“ und Abschaffung von Verbrennungsmotoren zu beheben. Das erste Problem ist für die natürliche Funktionsabläufe in einer Landschaft besonders gravierend. Es könnte zum Teil dadurch behoben werden, dass die Zerschneidungseffekte von Verkehrswegen durch grüne Brücken vermindert werden, noch effektiver durch großzügigen Brücken- und Tunnelbau. Dabei spielt die fachgerechte Ausführung und Unterhaltung der Grünverbindungen eine entscheidende Rolle (Peters-Ostenberg, Henneberg 2023).
Auch durch Alleen kann der schädliche Zerschneidungseffekt von Verkehrswegen gemindert werden. Außer ihrer Bedeutung als vernetzendes Element stellen sie selbst vielseitige Lebensräume dar.
Städte und Siedlungen
Zwischen 1985 und 2015 hat die die Ausdehnung von Städten und Siedlungen jährlich um 9687 km² zugenommen, mit steigender Tendenz (Liu et al. 2020). Damit ist der Flächenverbrauch der Städte schneller gewachsen als die Bevölkerung. Für eine nachhaltige Entwicklung müssen Städte deshalb „ökologischer“ werden. Damit ist gemeint, dass Funktionsabläufe in dem Ökosystem Stadt stärker den Funktionsabläufen in einem natürlichen Ökosystem entsprechen sollen. Eine Stadt mit großen Grünanlagen wie Parks und Gärten bietet zwar eine hohe Lebensqualität und eine bessere Ökobilanz. Dies geht aber insofern auf Kosten der Umgebung, als sie mehr Fläche für denselben umbauten Raum benötigt. Eine Erfolg versprechende Möglichkeit für dicht bebaute Großstädte ist die Integration von Bauwerken und Grünanlagen.
Neben Minderung des Klimawandels durch eine Verbesserung der CO2-Bilanz können dadurch auch die Auswirkungen einer Klimaerwärmung verringert werden (Lass u. a. 2022). Schließlich wirken mit Sachverstand begrünte Städte auch dem Verlust der Biodiversität entgegen.
Dächer
Schon lange zählt es zu Attributen ökologischer Bauweise, Dächer zu begrünen. Die Etablierung und Ausgestaltung solcher Dachgärten und Wiesen ist aber noch sehr stark ausbaufähig, wie man auf Luftbildern von Städten leicht erkennen kann. Begrünte Dächer können durch Brücken vernetzt werden. Durch treppenartige Anordnung von Gebäudeteilen können Verbindungen zur Grundfläche hergestellt werden. Beim Bewuchs selbst könnte dem Prinzip „Wachsen lassen“ mehr Raum gegeben werden.
Vernetzung von begrünten Dächern (Grafik W.Probst)
Fassaden
Auch begrünte Fassaden gibt es schon lange, aber eher an alten Bauernhäuser auf dem Land als an mehrgeschossigen Stadthäusern, Bankhochhäusern und Industrieanlagen. Außerdem sind die bisher architektonisch verwirklichten Grünfassaden gärtnerisch aufwändige Konstruktion, die eine hohe Wartung benötigen. Ziel müsste es sein, möglichst wartungsarme sich selbsterhaltende Systeme zu erzeugen.
Eine Möglichkeit für eine schnelle flächenhafte Begrünung wären Module, die mit einfachen Mitteln an Fassaden angebracht werden können und die durch Anschluss an eine Bewässerungsanlage wartungsarm sind. Die Elemente könnten aus einem Gerüst bestehen, an dem mehrere auswechselbare Pflanzgefäße aufgehängt werden. Fensterfassaden könnten durch berankte Schnurgerüste – Hopfenfeldern vergleichbar – begrünt und beschattet werden.
Fassadenbegrünung mit vorgefertigten Modulen (Grafik W. Probst)
Ein interessanter Vorschlag sind vorbegrünte Pflanzennetze. Solche „Urban Pergolas“ sollen als Verschattungssystem der Aufheizung von Fassaden entgegenwirken und die Städte in einen „diversen Großstadtdschungel“ verwandeln. Die Pflanzennetze können an einem oder zwischen mehreren Gebäuden angebracht werden und dadurch Grünflächen schaffen, ohne andere Nutzungen den Platz wegzunehmen (Urban Pergola 2021).
Balkone
Eine weitere Möglichkeit der vertikalen Begrünung, die in wenigen Beispielen schon verwirklicht ist, wäre die Ausgestaltung von Pflanzbalkonen mit Sträuchern und Bäumen (Boeri 2015).
Hochhäuser als Gewächshäuser, „Vertical Farming“
Diese platzsparende Form der Landwirtschaft setzt einen preisgünstigen Zugang zu alternativen Energien voraus, wird aber heute schon als eine wichtige, nachhaltige und zukunftsfähige Ergänzung zur Flächen gebundenen Landwirtschaft gesehen:
Wenn es in der Zukunft gelingt, den Kraftfahrzeugverkehr weitgehend aus den Stadtzentren herauszuhalten, werden dort auch keine Parkhäuser mehr benötigt und diese könnten zu „Plantscrapern“ werden (Despommier 2011).
Ritzen und Fugen
Der portugiesische Stadtplaner und Architekt Ángel Panero Pardo stellte auf dem großen Platz vor der Wallfahrtskathedrale von Santiago de Compostela während der Corona Pandemie fest, dass sich dieser Platz nach dem Ausbleiben der Pilger in ein Biotop für Wildkräuter verwandelt hatte. Die Fugen zwischen den Pflastersteinen waren grün. Der Stadtplaner überlegte, dass dieser zusätzliche Pflanzenwuchs sich eventuell positiv auf das Stadtklima auswirken könnte. Die Botaniker der Universität von Santiago de Compostela wurde mit einer Untersuchung beauftragt und sie stellten mit einer Wärmebildkamera fest, dass die bewachsenen Ritzen eine bis zu 28 °C niedrigere Oberflächen-temperatur aus aufwiesen als die Steine (Prinz 2023).
Dieses Ergebnis fand in den Medien einen breiten Widerhall, obwohl es eigentlich nicht so verwunderlich ist. Wenn man Fugen und Ritzen in Pflastern und Mauern nicht länger von jedem Bewuchs frei hält, sondern Bewuchs zulässt, hat dies einen messbar positiven Einfluss auf das Stadtklima.
Gehsteigkante mit Acker-Winde, Oberteuringen, 27.7.2016 (Foto W. Probst)
Gärten
Ein besonders großes Potenzial stellen Privatgärten dar, die vor allem in den Randbereichen der Städte in Vierteln mit Einfamilien- und Reihenhäusern konzentriert sind. Hier gilt meist das Prinzip, dass nur wachsen darf, was gepflanzt wurde. Der Garten darf nicht „verwildern“. „Un“kraut jäten ist deshalb neben Rasen mähen und Hecken schneiden die häufigste Beschäftigung des Hobbygärtners. Um das Unkraut ohne zu viel manuelle Tätigkeit fern zu halten, hat sich schon vor einigen Jahrzehnten verbreitet, die Beete mit einer Schicht aus keimungs- und wachstumshemmendem Rindenmulch zu bedecken.Seit einigen Jahren wird eine noch pflanzenfeindlichere Methode, das Auskiesen von Gartenflächen, immer beliebter.
Dabei gibt es viele Möglichkeiten, natürliche Funktionsabläufe im Garten zuzulassen oder sogar zu fördern und so eine „Verwilderung“ zu ermöglichen, die durchaus ästhetischen Ansprüchen gerecht werden kann:
Zierpflanzen, die gut gedeihen, fördern, auf solche, die schlecht wachsen oder sehr viel Pflege benötigen, verzichten,
auf Pestizide verzichten oder sie nur sehr gezielt bei einzelnen befallenen Pflanzen einsetzen,
Wildpflanzen nur entfernen, wenn sie gewünschte Zier- oder Nutzpflanzen schädigen oder verdrängen,
Wildpfanzen unter Hecken oder Sträuchern wachsen lassen,
Rasenflächen, die rein ornamentale Funktion haben, zu mageren (nicht gedüngten), höchstens zweimal im Jahr gemähten Wiesen umwandeln,
Abstellflächen (z.B. Autostellplätze) nicht pflastern oder asphaltieren, sondern als Schotterrasen gestalten,
Einfahrten mit unterbrochenen Pflastersteinen befestigen, die Bewuchs und Wasserversickerung ermöglichen,
abgeblühte Blütenstände und abgestorbene Fruchtstände wenigstens teilweise stehen lassen, auch über Herbst und Winter (Vogelfutter, Überwinterungsplätze für Insekten)
Gartenabfälle vor Ort kompostieren,
aus Strauch- und Baumschnitt Reisighaufen anlegen,
Gartenmauern als Trockenmauern anlegen, Mauerritzen können zur schnelleren Begrünung mit passenden Pflanzen geimpft werden (Zimbelkraut, Mauerraute, Schöllkraut, Polster von Mauermoosen wie Drehzahnmoos, Kissenmoos)
Abwechslungsreiche Besiedelungsflächen schaffen (Sandflächen, Lehmböden, humusreiche Böden, Stein- bzw. Bauschutthaufen),
Regenwasser vom Dach (und versiegelten Flächen) in Zisterne sammeln und als Gießwasser (ggf. auch für Teich /Bachlauf) nutzen.
Ergänzend zu den genannten Maßnahmen können Verkehrswege, insbesondere Straßen und Schienenverkehr, wie U-Bahnen unter die Oberfläche verlegt werden, wodurch Platz für bodenständige Grünanlagen aber auch Rad- und Fußwege gewonnen würde.
So könnten schließlich Städte entstehen, die ganz in einem grünen Pelz eingehüllt sind und die sich fast übergangslos in die umgebende Landschaft einfügen (vgl. Jean Nouvel 2014, Boeri 2015).
Verwilderung zulassen
Ein Garten, in dem verhältnismäßig wenig pflegerische Eingriffe vorgenommen werden, „verwildert“. Diese Art von Verwilderung ergibt sich aus natürlichen Funktionsabläufe, die nicht durch menschliche Eingriffe unterbrochen werden. Wenn man sich bei allen Eingriffen und Pflegemaßnahmen – Manipulationen der Natur – überlegt, welche Ziele mit Ihnen verfolgt werden sollen und ob diese Ziele notwendig und sinnvoll sind, wird man schnell erkennen, dass man auf viele Eingriffe verzichten könnte. Ein solcher Verzicht ist ein Schritt in Richtung Wildnis, wenn man unter Wildnis Vewilderung, das Zulassen natürlicher Prozesse, versteht.
Verwilderter Apfelgarten bei Flensburg (Foto U.Niss)
Quellen
Bibelriether, H. (2017): Natur Natur sein lassen. Die Entstehung des ersten Nationalparks Deutschlands: Der Nationalpark Bayerischer Wald. Freyung: Lichtland
Blanc.P. (2009): Vertikale Gärten, Die Natur in der Stadt. Stuttgart: Ulmer
Boeri, S. (2015): A vertical Forest. Milano: Editione Mantova
Bundesamt für Naturschutz: Neobiota – Gebietsfremde und invasive Arten in Deutschland. https://neobiota.bfn.de/
Crutzen, P. J. (2002): Geology of mankind. Nature 415, p.23
Daily, G. C. (2001): Ecological forecast. Nature 411, p.245
Despommier, D. (2011): The vertical farm: Feeding the world in the 21th century. Picador (Nachdruck der Ausgabe von 2010)
Fløjgaard, C. et al. (2021): Exploring a natural baseline for large-herbivore biomass in ecological restoration
Haft, J. (2023): Wildnis: Unser Traum von unberührter Natur (German Edition) (S.141). Penguin Verlag. Kindle-Version.
Hendersen, D. (o. J.): American Wilderness Philosophy. In: Internet Encyclopedia of Philosophy (IEP) http://www.iep.utm.edu/am-wild/
Kleunen, M. van et al. (2015): Global exchange and accumulation of non nativ plants. Nature 525, pp. 100–103
Lass, W., Reusswig, F, Walther, C.; Niebuhr, D.; Schürheck, T. Grewe, H. A. (2022): Hitzeaktionsplan für das Land Brandenburg (HAP BB). Gutachten, 20.9.22, Potsdam.
Liu, Xiaoping et al. (2020): High spatiotemporal resolution mapping of global urban change from 1985 to 2015: Nature Sustainability 3, pp.564-570.
Mantyka-Pringle, C. S., Martin, T. G., Rhodes, J. R. (2012): Interactions between climate and habitat loss effects on biodiversity: a systematic review and meta-analysis. Global Change Biology 18, pp. 1239-1252
MacArthur, R. H., Wilson, E. O (1967): The Theory of Island Biogeography. Princeton: University Press
Mancuso, S. (2021): Die Pflanzen und ihre Rechte. Eine Charta zur Erhaltung unserer Natur. Stuttgart: Klett-Cotta
Nickel et al. (2016): Außerordentliche Erfolge des zoologischen Artenschutzes durch extensive Ganzjahresbeweidung mit Rindern und Pferden: Ergebnisse zweier Pilotstudien an Zikaden in Thüringen, mit weiteren Ergebnissen zu Vögeln, Reptilien und Amphibien. Landschaftspflege und Naturschutz in Thüringen, 53, S. 5 – 20
Peters-Ostenberg, E., Henneberg, M. (2023): Zerschneidug – Entschneidung – Brücken bauen. In: : Riedel, W. (Hrsg., 2023): Zwischen Wildnis und Energielandschaft. Husum, S. 87-96
Sandom, C. J. et al. (2014): High herbivore density associated with vegetation diversity in interglacial ecosystems. In: Proceedings of the National Academy of Sciences of the United States of America, 111, 11, S. 4162 – 4167
Schilk,, D. (2019): Die Wiederbegrünung der Welt. Klein Jasedow: Drachen-Verlag
Tanneberger, F., Schroeder, V. (2023): Das Moor. München: dtv
Trommer, G. (2023): Der wilde Rest. In: Riedel, W. (Hrsg., 2023): Zwischen Wildnis und Energielandschaft. Husum, S. 195-218
Van Kolfschoten, T. (2000): The Eemian mammal fauna of central Europe. Netherlands Journal of Geosciences 79, 2,3, S. 269 – 281
Watson, J. E. M., Allen, J. A. u. a.: (2018): Protect the last of the wild. Nature 563, pp. 27-30
Weinzierl, H. (2007): Einführung zum Fachsymposium „Mehr Wildnis, die Zeit ist reif“. S. 6-8 in: Deutscher Naturschutzring (Hrsg.): Mehr Wildnis, die Zeit ist reif. Fachsymposium. Bonn
Wohlleben, P. (2013): Der Wald –ein Nachruf. München: Ludwig
Individuelle Fitness und natürliche Selektion sind zentrale Begriffe der Darwinschen Evolutionstheorie. In diesem Zusammenhang spielte der Begriff des „struggle for life“, des „Kampfes ums Dasein“, eine wichtige Rolle. Daraus wurde im Sozialdarwinismus nicht nur der „Sieg des Stärkeren“ sondern auch das „Recht des Stärkeren“ auf diesen Sieg als natürliches Recht abgeleitet. Dies wurde auch – nicht im Sinne Darwins, aber doch mit Bezug auf seine Theorie – auf die menschliche Gesellschaft übertragen und hatte starke Auswirkungen auf das Erziehungssystem, das lange Zeit auf die Förderung der Einzelleistung und die Qualifikation des Individuums abgestellt wurde.
Schon früh wurde diesen Folgerungen widersprochen, zum Beispiel von dem im englischen Exil lebenden russischen Anarchisten Pjotr Alexejewitsch Kropotkin (1842-1921) in seiner leider in Vergessenheit geratenen Schrift „Mutual Aid“. Auch die Erkenntnisse anderer Naturwissenschaftler zur großen Bedeutung der Kooperation bzw. des Mutualismus für die Evolution der Lebewesen gerieten in der ersten Hälfte des 20. Jahrhunderts in Vergessenheit. Eine Wiederbelebung erfuhren mutualistische Theorien erst wieder durch die bahnbrechenden Arbeiten Lynn Margulis` .
In der Wirtschaft hat Teamwork schon länger eine große Bedeutung. Gefördert durch die Digitalisierung und die elektronischen Medien hat sich in den letzten Jahrzehnten die Forderung nach gemeinschaftlichem Lernen in den Erziehungswissenschaften neu formiert unter den Begriffen „Kooperatives Lernen“ und „Kollaboratives Lernen“. Auch hier beruft man sich nicht nur auf Erkenntnisse der Pädagogik und der Sozielwissenschaften sondern auch auf die Biologie. Aus den Erkenntnissen über die Rolle der Kooperation bei der Evolution der Lebewesen leitet man ab, dass es eine genetische Disposition der Menschen für Kooperation und gemeinschaftliches Lernen geben muss (vgl. z. B. E.O. Wilson 2012).
Vor 20 Jahren, im Dezember 2010, erschien das von mir herausgegebene Unterricht Biologie Heft 280 „Miteinander“. Ich finde, angesichts dieser Entwicklungen ist es immer noch aktuell. Deshalb soll hier der Basisartikel im ungekürzten Entwurf mit einigen kleinen Korrekturen veröffentlicht werden:
Leben heißt Zusammenleben
„Don’t compete,combine!“ Kropotkin 1902
„Du sollst das Stroh zu Gold mir spinnen, ich brauche davon große Mengen“, fordert der Prinz die Müllerstochter auf und im Märchen bedient sich das arme Mädchen der besonderen Kenntnisse von Rumpelstilzchen. Auch wenn es wohl nie gelingen wird aus Gras Gold zu produzieren, so sind andere Transsubstantiationen doch fast genau so wunderbar: z.B. die Umwandlung von Gras in Milch, die von den Kühen in großem Maßstab für uns Menschen geleistet wird. Allerdings nicht von den Kühen alleine. Nur die Mikroben, in erster Linie die Bakterien, die in ihrem Pansen leben, schaffen es, den Hauptbestandteil des Grases, die Zellulose, aufzuschließen und damit der Verdauung und letzten Endes der Umwandlung in Milch zugänglich zu machen,
Herbivoren, Konsumenten erster Ordnung, sind wichtige Bestandteile von Ökosystemen, sie haben entscheidenden Anteil an der Regulation von Stoff- und Energiefluss, und sie alle sind dabei auf Verdauungshelfer in ihrem Darm angewiesen. Außerdem sind viele Herbivoren, wie die Vorfahren unserer Hausrinder, die Auerochsen, soziale Tiere, die in großen Herdenverbänden zusammenleben, in denen es Rangordnung und Kommunikation, gemeinschaftliche Aufzucht der Jungtiere und gegenseitige Hilfe gibt. Kein Wunder, dass gerade solche sozial lebenden Tiere vom Menschen domestiziert und genutzt wurden, eine neue Form der Wechselbeziehungen zweier Arten- durchaus nicht nur zum Vorteil des Menschen, mindestens, wenn man den genetischen Erfolg und die heutige Verbreitung der Haustierarten betrachtet. Bei der Verarbeitung der Milch schließlich bedient sich der Mensch wiederum mikrobieller Lebewesen, die man schon beinahe als „Hausmikroben“ bezeichnen könnte.
Schließlich gibt es einen weiteren Aspekt des neuen Miteinander von Menschen und Haustieren: Durch die Domestikation kamen neue infektiöse Keime in die menschlichen Populationen wie Pocken, Masern oder Influenza. Die Viehzüchter wurden dagegen allmählich immun, menschliche Populationen ohne Haustiere blieben sehr anfällig . Dies ist mit ein Grund für den raschen Niedergang indigener Kulturen nach der Kolonisation durch Europäer bzw. Asiaten (vgl. Diamond 1997)..
So gibt dieses Beispiel „Milch“ einen Eindruck von der Vielschichtigkeit des Miteinander des Lebens und der Lebewesen auf unserem Planeten.
Einmal zeichnet sich Leben durch Individualität und damit durch Grenzen und Grenzziehungen aus, zum anderen sind alle diese Grenzen – angefangen von den intrazellulären Membranen – „semipermeabel“. Wechselwirkungen über Barrieren hinweg sind ein Charakteristikum aller Lebensvorgänge und auch aller Lebewesen, insbesondere auch Wechselwirkungen mit anderen Lebewesen.
Bei der weiteren Darstellung dieser Wechselbeziehungen wollen wir sie zunächst in intraspezifische und interspezifische Beziehungen unterteilen, auch wenn dies gerade bei den Gruppen schwierig ist, bei denen sich – wie bei den Prokaryoten – Arten nicht eindeutig definieren lassen. Ein weiterer Abschnitt wird sich mit den Wechselbeziehungen innerhalb von Ökosystemen beschäftigen und schließlich soll die Rolle des Menschen als „hypersoziales Wesen“ in den Blick genommen werden.
Interspezifische Wechselbeziehungen
Forschungsarbeiten zum Thema „Symbiose“ bzw. „Kooperation“ haben lange Zeit eine relativ untergeordnete Rolle in der Biologie gespielt. Gerade im Hinblick auf die biologische Evolution wurde dieses wichtige Prinzip bis heute nicht ins rechte Licht gerückt – möglicherweise als Folge des unglücklichen Begriffs vom „Kampf ums Dasein“ und den mit diesem Schlagwort verbundenen populärwissenschaftlichen und biologistischen Interpretation der Evolutionstheorie (Sozialdarwinismus). Dabei gab es in der zweiten Hälfte des 19. Jahrhunderts durchaus wichtige Ansätze und die Zukunft weisende Erkenntnisse (vgl. Sapp 1994). Anton de Bary, ursprünglich Pflanzenpathaloge und Mykologe, definierte Symbiose als das Zusammenleben verschiedener Arten, wobei über den Nutzen für die einzelnen Arten noch nichts ausgesagt war. Er legte 1866 den Grundstein für die Erkenntnis, das Flechten einen Doppelorganismus aus Pilz und Alge darstellen. Der Begriff des Mutualismus wurde von dem Belgier Pierre-Joseph van Beneden 1873 geprägt (Vorlesung: Ein Wort zum Sozialleben Niederer Tiere). Albert Bernhard Frank – nach ihm sind die N2-assimilierenden Actinobakterien in den Erlenknöllchen „Frankia“ benannt – prägte 1877 den Begriff „Symbiotismus“ für alle Formen des engen Zusammenlebens verschiedener Arten ohne Berücksichtigung , wem dieses Zusammmenleben Vorteile oder Nachteile bringt. 1885 entdeckte er die Mykorrhiza. Der Berliner Karl Brandt und der Edinburgher Pattrick Geddes beschäftigten sich besonders mit „grünen“ niederen Tieren wie Hydra, Spongilla und Stentor. Geddes veröffentlichte 1881 einen Aufsatz über „Symbiosis of Algae and Animals“. Auch die Endosymbionten-Theorie der Eucyten wurde schon im letzten Jahrhundert geboren. A. F. W. Schimper äußerte 1883 als Erster die Vermutung, dass Plastiden Algen-Endosymbionten in Zellen sein könnten. Zu Beginn des 20. Jahrhunderts wurden diese Idee von den russischen Biologen Konstatine Sergejewitsch Mereschkowskii und Andrei Sergejewitsch Famintsyn aufgegriffen. Der von Famintsyn versuchte experimentelle Beweis – die isolierte in vitro Kultur von Plastiden und Mitochondrien – misslang allerdings. Die Theorie geriet zunächst in Vergessenheit. Erst in den letzten zwei Jahrzehnten des 20. Jahrhunderts wurde sie vollständig anerkannt (vgl. z.B. Margulis 1981, Schwemmler/ Schenk 1980). Es gilt heute als gesichert, dass es bei verschiedenen Algen zu mehrfachen Endosymbiosen (sekundären und tertiären Endosymbiosen) gekommen ist. Diese kamen dadurch zustande, dass eukaryotische fotosynthetisch aktive Algen durch Phagocytose aufgenommen und dann nicht vollständig verdaut wurden. Heute noch kann man diese Mehrfachendosymbiosen an der Zahl der die Chloroplasten umgebenden Membranen erkennen. Teilweise finden sich auch noch Kernreste in den Chromatophoren.
Die Beziehungen verschiedener Arten kann man – entsprechend der Einteilung in Kasten 1 – in Konkurrenz, Karpose (= Parabiose), Symbiose und Antibiose einteilen. Der Nutzen oder Schaden, den die Wechselbeziehung den Partnern gibt, kann durch Plus- und Minuszeichen bzw. durch eine Null bei Indifferenz ausgedrückt werden. Konkurrenz erhält dann zwei Minuszeichen, Karpose ein Plus und eine Null, Symbiose zwei Pluszeichen und Antibiose ein Plus- und ein Minuszeichen. Im Gegensatz zu dieser Einteilung fasste de Bary (1879) den Begriff der Symbiose weiter. Er verstand darunter einen Überbegriff für Parasitismus, Symbiose und Karopse, schloss allerdings Vereinigungen kurzer Dauer (z.B. das Zusammenwirken von bestäubenden Insekten und Blütenpflanzen) aus. Diese Symbiosedefinition hat sich vor allem im angelsächsischen Sprachraum bis heute erhalten. In der folgenden Darstellung halten wir uns jedoch an den hier gebräuchlicheren Einteilungsvorschlag im Kasten.
Tab. 1 Formen des Zusammenlebens zwischen verschiedenen Arten
Konkurrenzausschlussprinzip
Nach Hardin (1960) kann die Konkurrenzsituation zwischen zwei Arten kein Dauerzustand sein: Entweder wird eine Art verdrängt oder sie wandelt sich in ihren Ansprüchen und es kommt zu einem räumlichen (Allopatrie) oder ökologischen (Sympatrie) Nebeneinander. Dieses Konkurrenzausschlussprinzip ist eng gekoppelt mit der Definition der ökologischen Nische. Eine solche kann definitionsgemäß ebenfalls nur von einer Art gebildet werden. In Wirklichkeit sind die Verhältnisse allerdings etwas komplizierter. Dies hängt vor allem damit zusammen, dass es unter natürlichen Bedingungen keine Konstanz der Umweltfaktoren gibt. Im Laborversuch konnte Park 1954 und 1962 zeigen, dass zwei Mehlkäferarten mit nahezu identischen Umweltansprüchen sich unter Laborbedingungen gegenseitig verdrängen und dass es auf die Umweltbedingungen ankommt, ob die eine Art die andere oder die andere die eine verdrängt. Wenn aber unter natürlichen Bedingungen z.B. das Klima dauernd schwankt, so können auch zwei oder mehr Arten mit nahezu gleichen Ansprüchen in einem Lebensraum erhalten bleiben.
Symbiosen im engeren Sinne
Unter Symbiose in diesem Sinne versteht man sowohl langdauernde, eventuell sogar lebenslange enge Gemeinschaften, wie sie zum Beispiel zwischen den genannten Darmbakterien und den Rindern , zwischen Korallen und einzelligen Zooxanthellen oder auch zwischen Einsiedlerkrebs und Aktinie beschrieben werden als auch Gemeinschaften, die nur von kurzer Dauer sind. Hierzu gehören z.B. die Beziehungen zwischen blütenbestäubenden Insekten und Blüten (vgl. UB 236 Pflanzen und die sie bestäubenden Insekte). Andere Beziehungen – auch als „Allianz“ bezeichnet – kennzeichnen vorübergehende lockere Gemeinschaften wie die zwischen Madenhackern und Herbivoren oder zwischen großen Rifffischen und Putzerfischen.
Einige Beispiele:
Sehr charakteristisch sind Stoffwechselsymbiosen zwischen Tieren und Prokaryoten und Pilzen. Dies hängt damit zusammen, dass bei Pilzen und insbesondere auch bei Prokaryoten eine Vielzahl von Stoffwechselwegen entwickelt sind, die bei Eukaryoten fehlen. Durch die Symbiose können sich die Eukaryoten so Nahrungs- und Energiequellen erschließen, die anderenfalls verschlossen geblieben wären. Gleichzeitig profitieren die endosymbiotischen Mikroben von den Stoffwechselendprodukten ihrer Symbiosepartner und von den relativ geschützten Lebensräumen, die ihnen von diesen geboten werden.
Eine besonders enge Symbiose dieser Art besteht zwischen Blattläusen und den endocytosymbiotischen Bakterien der Gattung Buchnera, die in besonders großen Darmzellen (Bakteriocyten) leben. Die Endosymbionten werden von Generation zu Generation über die Eier weitergegeben. Buchnera ist eng verwandt mit Escherichia coli aber im Gegensatz zu diesem weit verbreiteten Darmbakterium ist ihr Genom wesentlich kleiner. Dafür sind in einer Zelle über 100 Kopien enthalten. Eine vollständige Genomanalyse von Buchnera ergab, dass keine Gene für Zelloberflächen-Lipopolysaccharide und Phospholipide vorhanden sind. Ebenso fehlen die meisten Regulatorgene und Gene, die der Verteidigung der Zellen nach außen dienen. Das enge Zusammenleben mit den Wirten wird dadurch deutlich, dass von Buchnera alle für seinen Wirt essentiellen Aminosäuren produziert werden. Dafür sind mindestens 55 Gene verantwortlich. Umgekehrt werden von dem Bakterium keine für den Wirt nicht essentiellen Aminosäuren produziert. Diese Komplementarität zeigt, dass die Symbiose schon sehr lange erfolgreich arbeitet. So bilden Blattläuse keine stickstoffhaltigen Exkrete, vielmehr produzieren sie Glutamin und dieses wird von den Bakterien als Ausgangsstoff für die Produktion von essentiellen Aminosäuren verwendet. Da Buchnera sogar seine Außenmembran vom Wirt erhält, kann man sagen, dass bei dieser Symbiose ein Stadium erreicht ist, das Buchnera schon fast als ein Zellorganell erscheinen lässt.
Noch komplizierter ist die Doppelendocytobiose in Darmzellen von Motten-Schildläusen. Wie durch Genanalysen nachgewiesen, enthalten die Bakterien in den Darmzellen ein weiteres endosymbiontisches Bakterium (von Dohlen, 2001: 433-436).
Viele ähnliche Beziehungen kommen bei Holz bzw. Zellulose fressenden Insekten wie Tabakskäfer, Borkenkäfer und Termiten vor. Auch blutsaugende Egel, Zecken und Läuse bessern die Inhaltsstoffe ihrer relativ einseitigen Nahrung durch symbiontische Darmbakterien auf. Sie können in besonderen Darmzellen (Bacteriocyten, auch Mycetome genannt), in Darmaussackungen oder auch frei im Darmlumen vorkommen.
Ein anderer Stoffwechsel-Symbiosetyp besteht zwischen Höheren Pflanzen und Prokaryoten, die das Luftstickstoffmolekül (N2) assimilieren können. Besonders bekannt sind diese Stickstoffendosymbionten der Gattung Rhizobium von den Hülsenfrüchtlern (Wurzelknöllchen). Bei anderen Höheren Pflanzen wie Erlen oder Sanddorn kommen stickstoffbindende endosymbiontische Aktinobakterien der Gattung Frankia vor. Schließlich können auch Blaugrüne Bakterien symbiontisch mit Höheren Pflanzen zusammenleben z.B. das Blaugrüne Bakterium Anabena azollae in besonderen Taschen des Schwimmfarns Azolla und andere Blaugrüne Bakterien in korallenartigen in den Luftraum ragenden Wurzeln von Cycadeen.
Noch nicht sehr lange bekannt sind die symbiotischen Beziehungen zwischen Sphagnen und methanotrophen Bakterien. Die Bakterien, die in den Wasserspeicherzellen der Sphagnen leben, nutzen die Oxidation des in tieferen Torfschichten gebildete Methan zur Energiegewinnung und sind gleichzeitig dazu in der Lage, Luftstickstoff zu assimilieren. Sie profitieren von der Sauerstoffproduktion und den teilweise abgegebenen Kohlenhydraten der Photosynthese betreibenden Sphagnum –Chlorocyten.
Blaugrüne Bakterien kommen auch als Symbiosepartner bei Flechten vor. Noch wichtiger sind bei dieser Symbiose aber eukaryotische Algen und zwar aus der Gruppe der Grünalgen, die mit Pilzarten eine sehr enge Gemeinschaft eingegangen sind. Hier ist durch die Symbiose eine völlig neue morphologische, ökologische und physiologische Einheit entstanden: Flechten sehen anders aus als die beiden Partner alleine, sie können völlig andere Lebensräume, auch extreme Standorte in der Arktis in Hochgebirgen oder in Wüsten, besiedeln und es gibt zahlreiche typische Flechteninhaltsstoffe wie z.B. Depside, Butenoide oder Azofarbstoffe, die jeweils vom einen der beiden Partner nicht gebildet werden können.
Außer in Flechten spielen Algen als Synbionten auch in vielen niederen Tieren eine bedeutende Rolle, so in den Polypen der Korallen, in marinen Würmern und Schnecken, in verschiedenen Muscheln und Schwämmen (vgl. UB 225 Algen, UB 254 Riffe). In allen diesen Fällen werden die heterotrophen Tiere durch die Algensymbionten teilweise autotroph. Bei einigen Strudelwürmern (z.B. Convoluta roskovensis) kann dies bis zum Verlust des eigenen Darmkanals gehen. Keeble (1910) hat hierfür die treffende Bezeichnung „Plant animals“ – Pflanzentiere – geprägt.
Eine weitere typische Symbiose zwischen Prokaryoten und Tieren stellen die verschiedenen Leuchtsymbiosen dar. Meerestiere wie Manteltiere, Tintenfische und Knochenfische nehmen die im Seewasser weit verbreiteten Leuchtbakterien in besonderen Organen als Symbionten auf und betreiben mit der Bakterienkolonie besondere Leuchtorgane.
Auch zwischen vielzelligen Tieren und Pflanzen gibt es zahlreiche symbiotische Gemeinschaften. Besonders bemerkenswert sind in diesem Zusammenhang die sogenannten Ameisenpflanzen oder Myrmecophyten. Sie stellen Hohlräume in Blättern, Stielen oder Wurzeln berei, die vor allem von Ameisen aber auch von anderen Kleintieren bewohnt werden. Besonders zahlreich kennt man solche Ameisenpflanzen aus Südostasien. Mehr als 150 Arten aus 27 Pflanzenfamilien wurden beschrieben.
Der baumförmige Schmetterlingsblütler Humboldtia laurifolia aus Sri Lanka hat hohle Internodien mit einem sich selbst öffnenden Eingang. Zusätzlich zu diesen „Wohnhöhlen“ (Domatium) bietet der Baum eine Reihe von extrafloralen Nektarien, die den Bewohnern gleichzeitig Futter liefern. Die Hohlräume werden vor allem von Ameisenarten besucht und besiedelt, besonders häufig von Technomyrmex albipes. Aber die Besiedelung kann auch durch verschiedene Ameisenarten erfolgen sogar in unmittelbarer Nachbarschaft. In einer gründlichen Untersuchung (Krombein et al. 1999) wurden weitere zehn Ameisenarten, verschiedene andere Insekten sowie Pseudoskorpione und Ringelwürmer als regelmäßige Bewohner der „Wohnhöhlen“ nachgewiesen. Bei einigen dieser Arten handelt es sich um eine sehr enge Gemeinschaft mit Humboldtia. Eine Vernichtung dieser Baum-Art würde auch zum Verschwinden der symbiontischen Bewohner führen.
Für die Pflanzen bringen die besiedelnden Ameisen vor allem einen Schutz gegen Fressfeinde. Die Wirksamkeit dieses Pflanzenschutzes wurde für das Symbiosepaar Crematogaster-Ameise und Ameisenpflanze Macaranga (Wolfsmilchgewächse) genauer untersucht. In diesem Falle werden von der Ameisenpflanze auch noch fett- und eiweißreiche Futterkörperchen für die Ameisen bereit gestellt. Die Ameisen revanchieren sich dafür, indem sie ständig die Oberfläche ihres Wirtsbaumes absuchen und diesen dabei von allem Fremdmaterial reinigen. Sie entfernen Insekteneier ebenso wie Raupen, Käfer und andere pflanzenfressende Gliedertiere. Auch pilzliche Krankheitserreger werden von den Ameisen beseitigt. Man konnte nachweisen, dass Macaranga mehr als 80 Prozent seiner Blattfläche verliert, wenn man die symbiontischen Ameisenkolonien entfernt (Linsenmaier, Heil 2001). Es konnte nachgewiesen werden, dass das Pflanzenhormon Jasmonsäure bei Macaranga die Blattnektarproduktion steuert (Boland et al.2001): Kommt es zu einer starken Schädigung durch Insektenfraß, wird die Hormonausschüttung erhöht und dies wiederum führt zu einer stärkeren Nektarproduktion. Dadurch lockt die Pflanze Ameisen, Wespen und andere Nektarkonsumenten an, die ihr helfen, sich gegen die Fraßfeinde zu verteidigen. Damit ist eine Form der Wechselwirkung zwischen Höheren Pflanzen angesprochen, deren Erforschung erst in jüngerer Zeit begonnen hat. Das Methyljasmonat ist einer von vielen Stoffen, der der Kommunikation zwischen Pflanzen dient. Pflanzen, die von Herbivoren z.B. Raupen befallen werden, produzieren in erhöhtem Maße Signalstoffe wie Jasmonat und diese bewirken bei anderen Pflanzen – auch bei anderen Pflanzenarten – eine verstärkte Produktion von für die Pflanzenfresser schädlichen Stoffen. Die Beziehung zwischen Blütenbestäubern und bestäubten Pflanzen ist in der Regel weniger eng. Es gibt jedoch auch hier Beispiele einer engen Gemeinschaft zweier Arten, die für beide lebensnotwendig ist. Dies gilt etwa für die komplizierten Wechselwirkung von Feige und Feigen-Gallwespe oder von Yucca und Yucca-Motte (Abb. ). Im natürlichen Lebensraum der Yucca-Arten im westlichen Nordamerika lebt ein kleiner Nachtfalter von etwa 13 mm Körperlänge, die Yucca-Motte (Pronuba yuccasella). Die Begattungsflüge der Yucca-Motten finden in der Dunkelheit statt. Das befruchtete Weibchen beginnt noch in der Nacht, die weißen duftenden Yuccablüten aufzusuchen und dort Pollen zu sammeln. Mit besonderen Fortsätzen der Kiefertaster wird der Pollen zu einem Klumpen geformt, der oft mehrere Millimeter Durchmesser haben kann und zwischen Kopf und Ansatz der Vorderbeine eingeklemmt und mit den Tentakeln seitlich festgehalten wird. Mit diesem Pollenpaket fliegt das Weibchen zu einer anderen Blüten derselben Art. Auf den dicken wachsachtigen Staubfäden sitzend, stößt es dann seine Legescheide durch die weiche Wand des Fruchtknotens in desse Höhlung hinein und legt an den Samenanlagen ein Ei ab. Sodann wandert das Tier entlang dem Stempel bis zur Narbe, stopft etwas von dem mitgebrachten Pollen in eine der drei Narbenfurchen oder in den dort offenen Griffelkanal, legt wieder ein Ei in den Fruchtknoten und so fort, bis sich eine Anzahl von Eiern im Inneren des Fruchtknotens befindet. Bald darauf wachsen die Pollenschläche von der Narbe durch den Griffelkanal zu den Samenanlagen und die Eizellen werden befruchtet. Während die Samenanlagen zum Samen heranwachsen, schlüpfen auch die jungen Räupchen aus den Eiern und sie beginnen, die heranwachsenden Samenanlagen aufzufressen. Nach einem Monat sind die Raupen ausgewachsen und verlassen den Fruchtknoten. Sie verpuppen sich in der Nähe der Yuccapflanze im Erdboden. Da die Raupen bis zum Verlassen des Fruchtknotens nur einen Teil der zahlreichen Samenanlagen verzehren, können sich viele noch zu reifen Samen entwickeln.
Antibiosen
Im Gegensatz zur Symbiose kennzeichnet die Antibiose eine Beziehung, die für einen der beiden Partner vorteilhaft, für den anderen aber schädigend ist. Ganz eindeutig gilt dies z.B. für die Beutegreifer-Beute-Beziehung . Nicht ganz so eindeutig ist dies für die für die Beziehung, die zwischen Pflanzen und Pflanzenfressern besteht, da die Konkurrenzkraft bestimmter Pflanzen durch regelmäßige Beweidung gestärkt wird. Auf solche Nahrungsketten soll jedoch hier nicht weiter eingegangen werden.
Demgegenüber bezeichnet man als Parasitismus, wenn ein Parasit einen Wirt ausnützt ohne dessen unmittelbaren Tod zu bewirken. De Bary definierte den Parasitismus folgendermaßen: „… der vollständige Parasitismus, d.h. jene Einrichtung, bei welcher ein Tier oder eine Pflanze den ganzen Vegetationsprozess durchmacht auf oder in einem anderen, einer ungleichnamigen Spezies angehörenden Organismus. Letzterer dient jenem, dem Parasiten, ausschließlich als Wohnort und liefert ihm sein gesamtes Nährstoffmaterial. Er ist in jeglichem Sinne des Wortes sein Wirt. Und jener lebt auf Kosten des Wirtes insofern sein Nährstoffmaterial die Lebendkörpersubstanz oder die zur eigenen Ernährung aufgenommene Nahrung dieses ist.“ Dabei weist schon De Bary darauf hin, dass es natürlich möglich sein wird, Parasiten auch außerhalb des Wirts künstlich am Leben zu erhalten, indem man ihnen eine geeignete Nährlösung bietet.
Eine für Parasiten besonders typische Erscheinung ist, dass sie oft auf mehrere Wirtsarten angewiesen sind, die sie im Laufe ihres Lebenszyklus sukzessive besiedeln. Häufig ist dieser Wirtswechsel auch noch mit einem Generationswechsel des Parasiten verbunden.
So gibt es vermutlich kaum eine höhere Pflanzenart, die nicht von einem oder mehreren Rostpilzen parasitiert wird. Eine große Zahl von Rostpilzen parasitieren auf Nutzpflanzen und sie sind deshalb für den Menschen von besonderer Bedeutung. Sicherlich ist dies ein Grund dafür, dass der Lebenszyklus vieler Rostpilze relativ gut erforscht ist (Gäumann 1959). Stellvertretend für die komplizierten Beziehungen der Rostpilze zu ihren Wirtspflanzen sei der Getreiderost (Puccinia graminis) erwähnt. Dieser Pflanzenparasit entwickelt sich einmal auf der Berberitze, zum anderen auf Getreidearten. Auf der Berberitze wächst das haploide Stadium des Basidiomyceten, auf der Getreidepflanze das Zweikernstadium. In überwinternden zweikernigen Dauersporen kommt es zur Kernverschmelzung und anschließend zur Meiose und zur Basidienbildung.
Nachdem der Entwicklungszyklus des gefährlichen Getriederostes aufgeklärt war, hat man in den 30er und 40er Jahren versucht, durch Ausrotten der Berberitze auch dem Rostpilz die Lebensgrundlagen zu entziehen. Dies gelang aber nicht, da in milden Wintern auch ungeschlechtlich produzierte Sporen überdauern und immer wieder zu einer Infektion der Getreidepflanzen führen können. Im übrigen werden solche Pflanzenparasiten immer durch große Monokulturen besonders gefährlich. Unter natürlichen Bedingungen können sich die Pflanzen gegen Parasiten sowohl tierlicher als auch pilzlicher Art recht gut verteidigen. Dabei kommt es teilweise auch zu einer Wechselwirkung zwischen recht verschiedenen Parasitenarten. So werden vom Verticillium-Pilz befallene Baumwollpflanzen weniger von parasitären Milben aufgesucht als nicht befallene und umgekehrt kann kein (kurzfristiger) Milbenbefall die Pflanzen resistenter gegen Pilzbefall machen (Martin 2002, S. 54/55).
Besonders zahlreiche Parasiten mit komplizierten Lebenszyklen kennt man vom Stamm der Plattwürmer (Plathelmintes). Typisch für die Saugwürmer (Trematoda), einer Klasse der Plathelminthes, ist eine endoparasitische Lebensweise in Darm, Leber, Lunge, Bindegewebe und Blutgefäßsystemen von Wirbeltieren. Sie haben einen relativ komplizierten Generationswechsel, der gleichzeitig mit einem Wirtswechsel verbunden ist: Aus den befruchteten Eiern der Tiere, die im Hauptwirt leben, schlüpfen in der Regel Wimpernlarven (Miracidien), die im ersten Zwischenwert zur Sporocyste werden. Aus der Sporocyste entstehen sogenannte Redien, die im zweiten Zwischenwirt zu Cercarien heranwachsen. Aus ihnen entwickeln sich, nachdem sie von Wirtstieren aufgenommen wurden, die adulten Geschlechtstiere. Bekannt ist das Beispiel des Kleinen Leberegels aus den Gallengängen von Schafen mit den Zwischenwirten Heideschnecke bzw. Zebraschnecke und Ameise. Das besondere an dieser Art von Parasitismus ist, dass der Parasit in diesem Fall die Ameise veranlasst, sich an Pflanzenstängeln festzukrallen. Dadurch wird sie besonders leicht von Schafen gefressen, was der weiteren Verbreitung des Parasiten dient. So abenteuerlich diese komplizierte, angepasste Lebensweise erscheint, so hat sich in jüngerer Zeit gezeigt, dass sie doch nicht einmalig ist. Ähnliche Erscheinungen kennt man von anderen parasitischen Trematoden: Microphallus piriformis lebt einmal in einer Strandschnecke (Littorina saxatilis), zum anderen in der Heringsmöwe. Es wurde nachgewiesen, dass von Trematoden befallene Strandschnecken die Tendenz haben, aufwärts zu kriechen, also in eine Position, in der sie leichter von Möwen gefressen werden können (MacCarthy 2000, 1161-1166). Befallene Schnecken verändern auch ihre Verhaltensweise bezüglich der Gezeiten. Im Gegensatz zu nichtbefallenen, kriechen sie gerade bei fallender Tide aufwärts. Nicht nur von Wirbellosen sondern sogar von Säugetieren kennt man eine solche parasitenbewirkte Verhaltensänderung: Ratten, die von dem Einzeller Toxoplasma gondii befallen sind, den sie vor allem aufsammeln, wenn sie Katzenkot fressen, werden neugieriger und weniger furchtsam. Das lässt sie zu einer leichteren Beute für Katzen werden und hilft so Toxoplasma, in seinen Hauptwirt zurückzukehren ( Berdoy 2000,1591-1594).
Karposen
Auf die große Zahl der Beziehungen, die für einen Partner mehr oder weniger vorteilhaft, für den anderen jedoch nicht schädigend sind, sei hier nur knapp eingegangen. Hierher gehören die Wohngemeinschaften (z.B. Fuchs und Brandgans) oder auch der zeitweilige Aufenthalt in Körperhöhlen von anderen Tieren. Die Nadelfische aus der Familie der Carapidae kommen mit etwa 25 Arten in wärmeren und warmtemperierten Meeren vor. Sie wohnen alle in Actinien, Seesternen, Seegurken, Feuerwalzen oder in Muscheln. Genauer wurde Carapus acus, ein mediterraner Nadelfisch untersucht. Er verlässt seien Wohnort Seegurke nur nachts. Um in die Seegurke hinein zu gelangen, schwimmt er mehr oder weniger senkrecht stehend mit wedelndem Schwanz um sie herum. Immer wieder wird dann der Versuch unternommen, am Hinterende in den Wirt einzudringen. Dazu stellt sich der Nadelfisch mit seinem Kopf dicht vor die Kloakenöffnung, führt seine Schwanzspitze am Körper entlang nach vorn und wahrscheinlich in dem Augenblick, in dem das Atemwasser in die Seegurke strömt, sich schnell umwendend, stößt er sein Hinterende in die Kloake. Dann dringt der Fisch nach und nach immer tiefer ein. Der Fisch dringt zunächst in die Wasserlunge der Seegurke ein, durchbricht diese aber dann, um sich in der Leibesöhle aufzuhalten. Während viele Nadelfische die Holothurien und andere Wirte nicht weiter schädigen, konnte man für Carapus acus nachweisen, dass er sich von den Geschlechtsdrüsen der Seegurke ernährt. Andere Arten kommen im Muscheln vor. Wenn die Fische in der Muschel sterben, werden sie als Fremdkörper mit einem Perlmuttüberzug versehen (z.B. Carapus homei aus der Karibik).
Auch die actinienbewohnenden Clownfische oder die als Muschelwächter bezeichneten Kurzschwanzkrebse, die in der Mantelhöhle von Muscheln zu finden sind, wären hier zu nennen. Die Aktinien könnten allerdings auch von den Futterresten der Clownfische profitieren, weshalb diese Partnerschaft oft auch als echte Symbiose bezeichnet wird.
Eine besonders große Rolle spielt die sogenannte Epökie (Aufsitzertum) – Lebewesen siedeln auf anderen. Besonders eindrücklich wird dieses Prinzip in den üppigsten Lebensräumen, wie Regenwäldern oder Korallenriffen, demonstriert. Aber auch bei uns gibt es viele „Epiphyten“ (Moose und Flechten auf Baumrinde) und „Epizoen“ (z.B. Seepocken auf Muschelschalen oder Krebspanzern, Glockentierchenkolonien auf Wasserflöhen und Ruderfußkrebsen.) Eine besondere Form der Wechselwirkung ist die der Transportgemeinschaft (Phoresie). Dungmilben und Fadenwürmer heften sich an Mistkäfer an, andere Milben werden von Weberknechten transportiert. Schiffshalter lassen sich von großen Fischen mitnehmen.
Intraspezifische Kooperation
„Wo ich auch immer das Tierleben in reicher Fülle auf engem Raum beobachtete, sah ich gegenseitige Hilfe und gegenseitige Unterstützung sich in einem Maße betätigen, dass ich in ihnen einen Faktor von größter Wichtigkeit für die Erhaltung des Lebens und jeder Spezies sowie ihrer Fortentwicklung zu ahnen begann.“ (Kropotkin 1902).
Artgenossen sind evolutionsbilogisch betrachtet von Natur aus Konkurrenten. Trotzdem kann man bei den Interaktionen zwischen Individuen einer Art, im allgemeinen auch als „Sozialverhalten“ bezeichnet, eine Vielzahl von Verhaltensweisen erkennen, die eindeutig koopertiv sind. Die ultimaten Ursachen solcher Verhaltensweisen sind z.B. die Fortpflanzung, die Brutpflege, die Fürsorge für die Jungtiere, der Schutz vor dem Gefressenwerden, der gemeinsame Beutefang oder die Sicherung von Weidegründen usw. Proximate Ursachen können in Erbkoordinationen oder in Lernvorgängen liegen.
Konkurrenz und Kooperation
Zu den Interaktionen zwischen Artgenossen gehören nicht nur kooperative Verhaltensweisen sondern auch Aggression und Konkurrenz, Rangordnung oder sogar Täuschung. Die Anpassungsselektion im Sinne Darwins ist, wenn auch nicht die einzige, so doch eine wichtige Grundlage der Evolution. Sie beruht auf dieser innerartlichen Konkurrenz und eventuell auf zwischenartlicher Kooperation. Die Individuen einer Art, die am meisten lebensfähige und überlebensfähige Nachkommen zeugen, geben damit ihre Gene und so auch viele ihrer Merkmale weiter. Der Kampf um einen Paarungspartner oder um eine Nahrungsressource ist häufig ritualisiert, d.h. für die Gegner besteht keine ernsthafte Verletzungsgefahr. Dies muss allerdings nicht so sein, wie man es z.B. von Kampfhähnen und Kampffischen weiß. Aber es ist einsehbar, dass bei Arten, bei denen die kämpfenden Rivalen sich verletzen , eine frühzeitige Beendigung des Kampfes die Fitness fördert, da sie auch den Sieger vor unnötigen Verletzungen schützt.
Unterschiedliches Balzverhalten ist eng gekoppelt mit unterschiedlichen Paarungssystemen. Während es bei vielen Tierarten keinerlei länger dauernde Paarbindungen gibt, kennt man andererseits monogame und polygame Beziehungen, wobei sowohl Polygynie als auch- seltener – Polyandrie vorkommen. Für die Paarungssysteme entscheidend dürften die besonderen Bedürfnisse der Jungen sein. So müssen Vögel nicht nur ihre Eier langwierig ausbrüten, die Jungen müssen dann auch mit erheblichen Nahrungsmengen gefüttert werden. Für beides ist eine Kooperation der Eltern von großem Vorteil. Deshalb ist eine monogame Paarbindung bei Vögeln häufig vorteilhaft – mindestens während der Brutzeiten und der Aufzucht der Jungen. Andererseits muss dies nicht unbedingt mit der rein monogamen Weitergabe der Gene, also mit der ausschließlichen Kopulation mit einem Geschlechtspartner, gekoppelt sein (vgl. Campell S. 1303, UB 185 Soziobiologie).
Häufig kommen in Tiersozietäten bestimmte Rangordnungen vor, die durch aggressive Auseinandersetzungen immer wieder gefestigt oder auch neu strukturiert werden. Für den reibungslosen Ablauf der innerartlichen Kooperation in einem Tierverband sind solche Rangordnungen u.U. von Vorteil, insbesondere wenn es um kompliziertes Zusammenarbeiten geht, wie etwa beim gemeinsamen Jagen. Bei Wölfen und anderen Hundeverwandten konnte nachgewiesen werden, dass die Rangordnung der weiblichen Tiere eines Rudels auch der Geburtenkontrolle dient: Wenn die Nahrung knapp ist, lassen die ranghöchsten Weibchen kaum Paarungen anderer, rangniedrigerer Weibchen zu. Sie sorgen damit dafür, dass nur ihre Gene weitergegeben werden. Ist reichlich Nahrung vorhanden, so lockern sie diese Restriktionen.
Schließlich ist das Revierverhalten ein wichtiger Bestandteil tierlichen Sozialverhaltens. Territorien oder Reviere dienen in der Regel der Sicherung der Nahrung, der Paarung und der Jungenaufzucht. Bei Vögeln werden solche Reviere häufig von Brutpaaren während der Brutzeit besetzt. Bei vielen Singvögeln müssen die Reviere relativ groß sein, weil sie auch der Nahrungsbeschaffung dienen. Bei Meeresvögeln können sie viel kleiner sein, da die Nahrung außerhalb des Reviers gesucht wird. Territorien werden häufig besonders markiert (Kot, Urin, Drüsensekrete). Auch akustische Markierungen wie lautes Brüllen der Seelöwen oder Gesänge der Singvögel dienen der Reviermarkierung. Territorialverhalten kann eine Population stabilisieren, da die Verteidigung des Territoriums verhindert, dass bei üppigem Nahrungsangebot eine Überpopulation entsteht, die dann u.U. wieder einen Zusammenbruch der Gesamtpopulation zur Folge hätte.
Altruismus
Altruismus oder uneigennützige Hilfeleistungen zwischen Individuen einer Art sind bei Tieren weit verbreitet. Besonders verbreitet sind Formen der Brutpflege, bei denen nur die Mütter, zum Teil Mütter und Väter und selten auch nur die Väter beteiligt sein können. Dabei geht es nicht nur darum, die Jungen zu füttern, zu wärmen und für ihr Wohlbefinden zu sorgen, sondern auch um die Verteidigung gegen Beutegreifer. Dies kann bis zur Aufoperfung des eigenen Lebens gehen. Die Soziobiologie versucht solche altruistischen Verhaltensweisen über die Fitness zu erklären. Bei der Brutpflege und bei der altruistischen Hilfe für Verwandte argumentiert die Soziobiologie mit dem Verwandtschaftskoeffizienten. Der Anteil der Gene, der bei zwei Individuen aufgrund gemeinsamer Abstammungen identisch ist. Der Verwandtschaftskoeffizient von Geschwistern beispielsweise beträgt 0,5, da 50 % der Gene von Geschwistern übereinstimmen. Für Cousins ersten Grades beträgt dieser Verwandtschaftskoeffizient 0,125. Es ist nach der soziobiologischen Theorie zu erwarten, dass sich Verwandte umso eher gegenseitig helfen, je höher dieser Koeffizient ist. Dies führt dann zu der sogenannten Familien- oder Verwandtschaftsselektion. (kin selection, Smith, Hamilton). Wenn man also sein Leben für zwei Kinder oder für acht Cousins opfert, so hat man genetisch bzw. evolutionsbiologisch gesehen, nichts verloren. Bei manchen altruistischen Verhaltensweisen ist eine Erklärung über den Verwandtschaftskoeffizienten allerdings nicht so eindeutig möglich. So warnen sich Murmeltiere gegenseitig durch Pfiffe vor Beutegreifern wie etwa Steinadlern. Wenn ein Adler oder ein anderer Fressfeind sich einer Murmeltierkolonie nähert, stößt eines der Murmeltiere einen schrillen Pfiff aus. Dadurch werden auch die anderen auf den Räuber aufmerksam und fliehen in ihre Baue. Das Pfeifverhalten allerdings wird für den Warner zu einem erhöhten Risiko. Nur wenn man davon ausgeht, dass die Murmeltiere einer Kolonie mehr oder weniger nahe verwandt sind, lässt sich dies ebenfalls über den Verwandtschaftskoeffizienten erklären. Eine andere Erklärungsmöglichkeit: Wenn alle Murmeltiere einer Kolonie zu diesem Verhalten bereit sind und es immer wieder ein anderes Tier trifft und dadurch der Nutzen und der Schaden ausgeglichen wird kann dieses Verhalten ebenfalls einen Fitnessgewinn bringen („Reziproker Altruismus“).
Tiergesellschaften
Die Vergesellschaftung von Individuen einer Art kann sehr unterschiedliche Organisationsmerkmale aufweisen. Unkoordinierte Verbände sind z.B. Schlafgemeinschaften, Überwinterungsgemeinschaften, Futtergemeinschaften. Ein gemeinsamer Ort und ein gemeinsames Ziel führen die Tiere zusammen. Man kennt solche Ansammlungen von vielen Insekten, aber auch bei Spinnentieren, Krebsen, Mollusken und bei allen Klassen der Wirbeltiere kommen sie vor. Solche Vergesellschaftungen sind in der Regel zeitlich begrenzt, die einzelnen Individuen können sich leicht wieder von der Gruppe lösen, und die Koordination zwischen den einzelnen Individuen ist gering. Schon etwas anders sieht es bei koordinierten Verbänden aus, wie sie etwa bei ziehenden Vogelschwärmen, wandernden Libellen, Heuschrecken und Schmetterlingen oder Fischschwärmen vorliegen. Hier findet oft eine erstaunliche Koordination der Flug- oder Schwimmbewegungen statt. Der soziale Gesichtspunkt ist deutlich, gemeinsamer Aufbruch zur Wanderung, gemeinsame Bestimmung des Zieles oder Lösung des Orientierungsproblemes. Gefahr von außen, etwa ein herabstürzender Raubvogel auf einen Starenschwarm, führt zu koordinierten Reaktionen. Der Schwarm kondensiert sich, bildet einen Stoßpulk, der gemeinsam zum Angriff übergehen kann. Anders reagiert ein Elritzenschwarm, bei dem ein Mitglied vom Hecht ergriffen wurde. Der ganze ergreift panikartig die Flucht, was damit zusammenhängt, dass von der Bißwunde der verletzten Elritze ein hochwirksamer Schreckstoff freigesetzt wird. Die so gewarnten Elritzen meiden den Ort, an dem ihr Genosse gefressen wurde, wochenlang. Hier handelt es sich also um ein soziales Warnsignal, das in Hautzellen gespeichert wird und dass ohne jeden biologischen Nutzen für das individuelle Leben ist, das aber für den Gesamtverband große Vorteile bringt.
Andere Schutzgemeinschaften finden sich z.B. bei verschiedenen Insekten. So können sich Feuerwanzen zu größeren Verteidigungsgemeinschaften zusammenschließen. Sie besitzen Verteidigungsdrüsen und damit verbunden eine Warnfärbung. Die Warnwirkung wird durch die Gruppierung erhöht.
Soziale Verbände höherer Organisation sind charakteristisch für die Insektenstaaten. Hier ist die soziale Bindung obligatorisch. Der Verband bildet eine geschlossene Gemeinschaft und es kommt zu einer Differenzierung der Individuen. Sämtliche Entwicklungsstadien von Eiern, Larven über Puppen bis zu den geschlechtsreifen Imagines sind Bestandteil dieses Verbandes. Alle Tätigkeiten, die für den Fortbestand für die Gemeinschaft wesentlich sind, werden im Kollektiv und arbeitsteilig ausgeführt. Häufig kommt es zu einer Kastenbildung, d.h. die ausgewachsenen Tiere haben – entsprechend ihren unterschiedlichen Aufgaben einen unterschiedlichen Körperbau. Typisch für das Funktionieren solcher Tierstaaten ist eine meist angeborene, sehr differenzierte Fähigkeit zur Kommunikation. Teilweise werden solche Staten als „Überorganismen“ bezeichnet.
Termiten z.B. sind besonders hoch organisierte staatenbildende Insekten mit mindestens drei Kasten: Königin und König als Geschlechtstiere leben immer zusammen, außerdem werden Arbeiter und Soldaten oft noch in verschiedenen Ausprägungen ausgebildet. Soldaten und Geschlechtstiere können sich nicht selbständig ernähren und sind auf die Fütterung durch die Arbeiter angewiesen. Insgesamt beruht das Zusammenspiel innerhalb des Termitenstaates auf komplizierter Kommunikation, teilweise auf der Basis von Pheromonen. Außerdem leben Termiten auch noch mit anderen Lebewesen in Symbiose. Als Pflanzenfresser können sie mit Hilfe von endosymbiontischen Bakterien und Einzellern Zellulose zersetzen. Andere Arten können sogar den Ligninstoff mit Hilfe von Pilzendosymbionten aufschließen. Außerdem werden von Termiten Pilzgärten angelegt, und zwar auf einem Gemisch aus Kot und zerkauter Nahrung und Holz. Die Pilzgeflechte dienen vor allem den Larven als Nahrung.
Die Zusammenarbeit im Termitenstaat sorgt für gleichbleibende Innentemperaturen, günstige Feuchtigkeitsbedingungen und sichere Aufzucht der Nachkommen (vgl. UB 169, S. 45).
Im Prinzip ähnlich, aber meist nicht ganz so kompliziert, sind die Verhältnisse bei Ameisen und bei anderen Hautflüglern. Bemerkenswert ist die besonders komplizierte Form der Kommunikation der Honigbienen (vgl. Hedewig 2000 in UB 260).
Symbiose in Ökosystemen
Für das Wirkungsgefüge eines Ökosystems bilden symbiotische Beziehungen eine entscheidende Rolle, obwohl sie bisher in der ökologischen Literatur noch relativ wenig berücksichtigt wurden. Ganz allgemein kann man sagen, dass Ökosysteme dazu tendieren, im Laufe ihrer Entwicklung an Komplexität zuzunehmen. So sind die ältesten Ökosysteme gleichzeitig die komplexesten und die Wechselbeziehungen solcher Systeme sind besonders kompliziert. Hier soll etwas ausführlicher auf die Bedeutung der Symbiosen für den Stoffkreislauf in Ökosystemen eingegangen werden. Da die Chloroplasten aller Höheren Pflanzen und Algen aus endosymbiontischen Prokaryoten hervorgegangen sind, wird der größte Teil der Primärproduktion in der Biosphäre durch eine Symbiose geleistet. Abgesehen davon spielen Flechten als Primärproduzenten auf etwa einem Achtel der Landfläche (1,2 x 107 km2) eine entscheidende Rolle. Bedeutend ist weiterhin der Beitrag der Korallenriffe zur Primärproduktion.
Die enge Partnerschaft von Pilzen und Landpflanzen besteht vermutlich seit der Eroberung des Landes im ausgehenden Silur. Für Primärproduktion und Stoffkreislauf in der Biosphäre ist diese Symbiose von Höheren Pflanzen und Pilzen (Mykorrhiza) besonders wichtig. Man nimmt an, dass Mykorrhizapilze 10 bis 20% der fotosynthetischen Primärproduktion von Pflanzen aufnehmen, das sind bis zu 2 x1013 kg pro Jahr. Besondere Bedeutung haben Mykorrhiza-Pilze für de Phosphor- und Stickstoff-Kreislauf in Ökosystemen . (vgl. Agerer, UB ), Sie stellen eine Kurzschluss artige Verbindung zwischen organischen Abfallstoffen und Primärproduzenten her. Dadurch kann der Export dieser Elemente aus Ökosystemen deutlich verringert werden. Wie Perakis und Hedin (2002, S. 416-418) nachweisen konnten, ist der Austrag an anorganischem Stickstoff in naturnahen, vom Menschen wenig beeinflussten Regenwäldern des gemäßigten Südamerika viel geringer als in entsprechenden, stark vom Menschen beeinflussten Wäldern Nordamerikas (Abb. ) . Dies könnte darauf hindeuten, dass es in diesen Wäldern kaum zu einer totalen Remineralisierung von Stickstoffverbindungen kommt. Ähnliches dürfte für Phosphorverbindungen gelten.
Die Verbreitung von Mykorrhizapilzen ist viel größer, als man dies ursprünglich angenommen hatte. So gilt als sicher, dass in tropischen Wäldern etwa 90 % aller Gehölze Mykorrhizen ausbilden. Da viele Pilze mit mehreren Baumarten Partnerschaften eingehen, ist nicht nur ein Stofftransport von einem Baum zum anderen sondern auch von einer Baumart zur anderen möglich. Besonders spektakulär ist in diesem Zusammenhang, dass durch Pilzwurzeln vermittelt auch Pflanzen existieren können, die kein Chlorophyll mehr bilden und dann als reine Parasiten auf den Pilzen leben. So findet man das bleiche Wintergrüngewächs Fichtenspargel unter Fichten und Buchen. Vermittelt durch einen Mykorrhizapilz lebt der Fichtenspargel von der Primärproduktion der Buchen bzw. Fichten.
Schließlich können Pilze auch tierische Eiweißquellen für Höhere Pflanzen erschließen, die sonst nur den Extremspezialisten – den sogenannten Carnivoren oder Insektivoren – vorbehalten bleiben. So konnte nachgewiesen werden, dass Laccaria bicolor (Zweifarbiger Lacktrichterling) Springschwänze (z.B. der Art Folsomia candida) „fressen“ kann. Der Pilz immobilisiert die Springschwänze zunächst. Dann dringt das Mycel in deren Körper ein und fängt an, sie zu „verdauen“. Da Laccaria gleichzeitig mit Waldbäumen eine Mykorrhiza eingeht, werden tierliche Stickstoffverbindungen über den Pilz an die Bäume weitergegeben. Durch 15N-Isotopenmarkierung konnte nachgewiesen werden, dass bis zu 25 Prozent des pflanzlichen Stickstoffs aus Springschwänzen stammen, die von Laccaria gefressen wurden. Als Gegenleistung versorgt der Baum den Pilzpartner mit Kohlenhydraten, auch mit solchen, die dann zu proteolytischen Enzymen umgebaut werden können (Klironomos, Hart 2001,p.651,652).
Untersuchungen an isolierten Rasenstücken ergaben, dass durch den von Mykorrhizapilzen vermittelten Stoffaustausch die Konkurrenz zwischen den Pflanzenarten vermindert wird. Dies führt dazu, dass die Zugangsmöglichkeiten zu Mineralstoffen ausgeglichen werden und dass die Koexistenz verschiedener Arten leichter ist. Eine Zunahme der Artenvielfalt ist die Folge (A.E. Douglas Symbiotic interactions 1994).
Auch im Zusammenhang mit Primärsukzessionen (Neubesiedelung von vorher vegetations- bzw. organismenfreien Substraten etwa nach Vulkanausbrüchen oder beim Gletscherrückzug) können Symbiosen eine wichtige Rolle spielen. Dies gilt etwa für die Neubesiedelung von Gletschern freigegebener Felsflächen durch Flechten und anschließend durch Pflanzen, die in Symbiose mit Luftsticksoff – fixierenden Bakterien leben. Genauere Untersuchungen in Glacer Bay/ Alaska, wo ein Gletscher sich in überlieferten Zeiträumen um etwa 100 km zurückgezogen hat, zeigt eine ständige Zunahme von Stickstoff in den Böden. Als Erstbesiedler an Höheren Pflanzen spielt die Silberwurz (Dryas) eine entscheidende Rolle. Sie enthält stickstofffixierende Bakterien der Gattung Frankia. In der Folgezeit besiedeln Erlen, Weiden und Pappeln die Gletscherrückzugsgebiete. Die endosymbiontischen Actinomyceten der Gattung Frankia können in Erlenbeständen bis zu 180 kg Stickstoff/ ha und Jahr fixieren (Abb. Grafik zur Stickstoffzunahme in Gletscherböden von Glacer Bay). In bestimmten limnischen Lebensräumen, z.B. in ostasiatischen Reisfeldern, dürfte die Stickstofffixierung durch mit dem Schwimmfarn Azolla zusammenlebenden Blaugrünen Bakterien der Gattung Anabena eine wichtige Rolle spielen (50 bis 150 kg Stickstoff/ ha und Jahr). In tropisch-subtropischen marinen Lebensräumen des pazifischen Raumes spielt die Diatomee Rhizosolenia mit endosymbiontischen Blaugrünen Bakterien eine ähnlich bedeutende Rolle beim Zugang des Ökosystems zur Luftstickstoffquelle.
Erst in jüngster Zeit beginnt man, die komplizierten Wechselwirkungen zu studieren, die zwischen den Mikroorganismen und dem höheren Leben der Ozeane bestehen. Die Bedeutung der Prokaryoten und insbesondere der ursprünglichen „Domäne“ der Archaea ist mengen- und massenmäßig in den Ozeanen viel bedeutender als lange Zeit angenommen. So ist reiches Archaea-Vorkommenie keineswegs auf die Umgebung der schwefelspuckenden Tiefseeschlote begrenzt. Vielmehr dürften sie 40 % der Tiefseeorganismen insgesamt ausmachen und die Tiefsee ist bei weitem der größte Lebensraum der Erde. Aber auch in oberflächennahen Wasserschichten der Ozeane spielen Archaebakterien eine große Rolle. Die zur Fotosynthese fähigen a-Proteobacteria machen vemutlich wenigstens 10 % aller Bakterien in den Ozeanen aus (Copley 2002) und sie sind damit für ca. 5 % der Fotosynthese bedingten Primärproduktion verantwortlich. Dabei läuft ihre Fotosynthese allerdings etwas anders ab. Sie produzieren nämlich keinen Sauerstoff, sondern verwerten den bei der Fotosynthese freigesetzten Sauerstoff sofort wieder für eigene Synthesen. Außerdem nutzen sie Lichtenergie, um organische Verbindungen abzubauen (Fotoheterotrophie). Diese Fähigkeit, von der man bis vor kurzem nichts wusste, hat bedeutende Auswirkungen für den Kohlenstoffkreislauf und die mögliche Bedeutung der Ozeane als Kohlenstoffsenke. Ein großer Teil des Kohlenstoffdioxids, das in den Ozeanen durch die Fotosynthese fixiert wird, bleibt – wenn die Planktonorganismen absterben – in gelöstem oder suspendiertem organischem Material zurück. Dieses organische Material dürfte in größerer Menge als bisher angenommen von Proteobakterien genutzt werden. So tragen diese Prokaryoten dazu bei, dass der Kohlenstoff stärker als bisher angenommen in den oberen Wasserschichten bleibt – eine schlechte Nachricht für diejenigen, die bisher hofften, dass übermäßiger Anstieg des Kohlenstoffdioxidgehaltes der Atmosphäre durch die Ozeane ausgeglichen werden kann. Wie neue Arbeiten zeigen, gibt es noch eine große Zahl anderer Prokaryoten mit außergewöhnlichen Stoffwechsel- und Fotosynthesewegen, die bisher noch nicht erforscht sind und die eine Vielzahl neuer Beziehungen und Stoffflussschleifen erwarten lassen, die das Zusammenspiel in dem von Planktonorganismen bestimmten Ökosystem der freien Ozeane viel komplizierter erscheinen lassen wird, als dies bisher angenommen wurde. So dürfte die von Halobakterien als Fotergie bekannte Erscheinung, bei der Sonnenlicht über Rhodopsin und verwandte Pigmente als Energiequelle für eine Membran-Protonenpumpe genutzt wird, in oberflächennahen marinen Habitaten weit verbreitet sein. Auch die Bedeutung Stickstoff-(N2)-fixierender Prokaryoten in marinen Ökosystemen ist vermutlich unterschätzt worden. So dürfte insbesondere die Zahl der entocytosymbiotischen Stickstofffixierer in den einzelligen Planktonalgen eine viel größere Rolle spielen als bisher angenommen (Zehr 1998). Auch Viren sind häufig in marinen Ökosystemen. Sie können bei Algenblüten regulierend wirken. Wenn die virusbefallenen Zellen solcher großen Algenblütengebiete plötzlich sterben, setzen sie eine große Menge organischer Materie frei. Auf diese Art und Weise können Viren möglicherweise zur plötzlichen massenhaften Freisetzung von Dimethylsulfid (DMS) beitragen. Wie zum ersten Mal von Lovelock nachgewiesen, fördert DMS in der Atmosphäre die Wolkenbildung und erhöht damit die Menge der an der Atmosphäre reflektierten Sonnenstrahlen. Dies kann eine deutliche Abkühlung des Erdklimas bedeuten. Es ist durchaus naheliegend, anzunehmen, dass die große Stabilität der Biosphäre solchen mikrobiellen Wechselwirkungen zu verdanken ist. Im Kleinen kennt man solche eng kooperierende Mikrobensysteme, in denen sich die einzelnen Bestandteile gegenseitig stabilisieren, z.B. von denen von Kefir oder Kombucha.
Eine besondere Form der Wechselwirkung ist der durch Viren vermittelte horizontale Gentransfer. Auch diese Form der Wechselwirkung wurde vermutlich in der Vergangenheit eher unterschätzt.
Menschliche Macht, Mitgefühl und Zukunftsfähigkeit
Je höher entwickelt das Nervensystem und das Lern- und Erinnerungsvermögen einer Tierart, desto flexibler und anpassungsfähig kann auch das Sozialverhalten werden. Junge werden „geprägt“, Kinder lernen von ihren Eltern, schließlich können sich sogar Traditionen herausbilden. An der Spitze dieser Entwicklung stehen zweifellos die Primaten und schließlich die Menschen.
Sicherlich wirkt sich dies auch auf das Kooperationsverhalten aus. So konnte durch Computersimulationen nachgewiesen werden, dass kooperatives Verhalten den kooperierenden Individuen Selektionsvorteile bringt, wenn die Individuen innerhalb einer Population erkennen können, ob andere Individuen kooperationsbereit sind oder nicht. Dies setzt ein hochdifferenziertes Wahrnehmungsvermögen voraus. (Sigmund, Nowak 2001, 403,404).
Auch die sogenannte Altruistische Bestrafung (Altruistic punishment, Fehr, Gächter, 2002, 137-140) stellt eine Verhaltensweise dar, die Altruismus fördert: Sie beschreibt ein typisch menschliches Verhalten gegenüber Individuen, die agressiv ihre eigenen Interessen verfolgen. Solche Individuen werden bestraft, auch wenn der Strafende davon keinen direkten eigenen Vorteil hat. Diese Verhaltensweise setzt voraus, dass man sich relativ gut in andere Individuen hineinversetzen kann – eine Fähigkeit die Menschen in höherem Maße haben als andere Arten.
Wahrnehmungsfähigkeit und Gehirn gestatten es den Menschen, sich ein Bild ihrer Umwelt zu machen, das planvolles und gezieltes Handeln erlaubt. Die abstrakte Sprache macht es möglich, sich intensiv und detailliert mit anderen Menschen auszutauschen, Erkenntnisse, Einsichten, Erfahrungen und Ideen weiterzugeben. Die Schrift und seit Kurzem die elektronischen Medien erlauben eine Konservierung von Information und in Zukunft vielleicht auch eine enge Kooperation von Gehirnen und Systemen der elektronischen Datenverarbeitung. So wird von manchen Informatikern eine Symbiose zwischen menschlichem Gehirn und Maschine angedacht: „Schließlich werden wir darangehen die externen Hilfsmodule mit dem Gehirn zu verbinden – beispielsweise durch Millionen mikroskopischer Elektroden; man könnte sie in das große Faserbündel namens Corpus callosum implantieren, das als gewaltiger Datenbus die beiden Gehirnhälften miteinander verbindet.“ (Minsky 1994).
Mit Hilfe seines Gehirns kann der Mensch Szenarien in Gedanken durchspielen, die Folgen bestimmter Handlungen vorhersehen. Dadurch, dass Beziehungen und Wechselwirkungen in die Zukunft projiziert werden, kann man ihre Folgen abschätzen. Diese Modellbildungsfähigkeiten sind ein enormer Machtfaktor. Menschen neigen allerdings dazu, diese Macht zu überschätzen und damit gleichzeitig die Verantwortung zu unterschätzen, die aus der Erkenntnisfähigkeit erwächst. Denn die Modelle, die eben doch nur Modelle und damit unvollkommene Bilder der Wirklichkeit sind, werden oft als ganz real genommen und man verlässt sich voll auf ihre Aussagen. So werden mit der Zunahme menschlicher Macht und Manipulationsmöglichkeiten die als Folgen menschlicher Eingriffe auftretenden Katastrophen immer größer und gefährlicher.
Vielleicht hilft ein Nebeneffekt unseres Weltbildapparates dabei, das richtige Maß zu finden: Er gestattet es den Menschen nämlich auch, sich in andere Individuen hineinzuversetzen, die Welt „mit ihren Augen“ zu sehen. Diese Fähigkeit ist Voraussetzung für Mitgefühl, Mitleid und die Möglichkeit, sich mit Anderen zusammen zu freuen und damit die Voraussetzung für Wertempfinden, Ethik und Moral.
Dabei betrifft Mitfühlen und Mitleiden nicht nur Mitglieder der eigenen Art, sondern auch andere Mitgeschöpfe. In ihrem Versuch „Leben“ zu beschreiben und verständlich zu machen weisen Lynn Margulis und Dorian Sagan besonders auf dieses menschliche Bewusstsein hin, das im Grunde ein Bewusstsein der Biosphäre oder des gesamten Lebens der Erde darstellt, da es allmählich mit der Evolution gewachsen ist. „In diesem Sinne ist das intuitive Wissen und Werden, nach dem jedes Einzelbewusstsein eine Illusion ist und wir alle einem einzigen Urgrund, nämlich Brahman angehören, vielleicht völlig richtig: Nicht nur unsere chemische Zusammensetzung ist ein gemeinsames Erbe, sondern auch unser Bewusstsein und die Notwendigkeit in einem Kosmos zu überleben, der aus der gleichen Materie besteht wie wir, der aber unserem Leben und unseren Belangen gleichgültig gegenübersteht.“ (Margulis, Sagan 1997).
Literatur
Agerer, R.: Mykorrhiza. UB 183/17:49-51, 1993
Ahmadjian, V., Paracer, S.: Symbiosis: An introduction to biological associations. Univ.Press New England, Hanover (USA) 1986
Barlow, C. (Hrsg.): Evolution extended. Biological debates on the meening of life. MIT Cambridge (USA), London, 1994
Berdoy,M, Webster, J.P., Macdonald, D.W.: Fatal attraction in rats infected with Toxoplasma gondii: Proceedings of the Royal Society of London B267:1591-1594, 2000
De Bary, A.: Die Erscheinung der Symbiose. Vortrag gehalten auf der Versammlung Deutscher Naturforscher und Ärzte zu Cassel. Trübner, Strasburg 1879
Copley, J: All at sea. Nature 415, 7.2.2002: 572-574
Diamond, J.: Guns, Germs and Steel. Norton, London, New York 1997
Krombein, K. V. et al.:Biodiversity of domatia occupants (ants, wasps, bees and others) of the Sri Lankan myrmecophyte Humboldtia laurifolia VAHL (Fabaceae).Smithonian Contributions to Zoology 603, Washington (USA) 1999
Kropotkin, P.: Gegenseitige Hilfe in der Tier- und Menschenwelt. Trotzdem-Verlag, Glasenau 19932 (Übersetzung der englischen Originalausgabe von 1902: Mutual aid)
Margaris, N.S., Arianoustou-Faraggitaki, M., Oechel, W.C. (Hrsg.): Beeing alive on land. Junk Publishers, The Hague, Boston, Lancaster 1984
Margulis, L. (Hrsg.): Symbiosis in cell evolution 1981; 2nd ed.: Endosymbiosis in cell evolution. Freeman, San Fransisco (USA) 1993
Margulis, L., Fester, R. (Hrsg.): Symbiosis as a source of evolutionary innovation: Speciation and morphogenisis. MIT Cambridge(USA) 1989
Margulis, L., Sagan, D.: Leben. Vom Ursprung zur Vielfalt. Spektrum, Heidelberg, Berlin 1999
Masuch, G.: Biologie der Flechten. Quelle & Meyer, Heidelberg, Wiesbaden 1993
Martin, K.: Ökologie der Biozönosen. Springer, Berlin, Heidelberg, New York 2002
McCarthy, H.O., Fitzpatrick, S., Irwin, S. W. B.: A transmissible trematode affects the direction and rhythm of movement in a marine gastropode. Animal Behavior, London 59: 1161-1166, 2000
Minsky, M.: Werden Roboter die Erde beherrschen? In: Spektrum der Wissenschaften, Spezial 3: Leben und Kosmos, S.80-87, Heidelberg 1994
Perakis, S.S., Hedin, L.O.: Fluxes and ftes of nitrogen in soil of an unpolluted old-growth temperate forest, southern Chile. Ecology 82(8):2245-2260, 2001
Probst, W. (Hrsg.) :Algen. UB 225/21, 1997
Probst, W. (Hrsg.) Pflanzen und Insekten. UB 236/22, 1998
Die Werte, auf die sich die Union gründet, sind die Achtung der Menschenwürde, Freiheit, Demokratie, Gleichheit, Rechtsstaatlichkeit und die Wahrung der Menschenrechte einschließlich der Rechte der Personen, die Minderheiten angehören. Diese Werte sind allen Mitgliedstaaten in einer Gesellschaft gemeinsam, die sich durch Pluralismus, Nichtdiskriminierung, Toleranz, Gerechtigkeit, Solidarität und die Gleichheit von Frauen und Männern auszeichnet. (Artikel 2 EUV in der Fassung des Vertrags von Lissabon 2009)
Im Zusammenhang mit dem Ukraine-Krieg wird häufig betont, dass es hier um die Verteidigung „europäischer Werte“ gehe. Diese allgemeine Formulierung ist höchst missverständlich, denn die Werte, die in den Gesellschaften der europäischen Staaten über Jahrhunderte galten, stimmen nicht mit den Grundwerten der Europäischen Union überein, die diese sich zuletzt in dem Vertrag von Lissabon 2009 gegeben hat. Ganz im Gegenteil! Es wäre deshalb wesentlich zielführender und treffender, von „humanistischen Werten“ zu sprechen, deren Quellen keineswegs auf Europa beschränkt sind und die von den meisten europäischen Staaten seit Jahrhunderten mit Füßen getreten werden.
Besonders deutlich wird dies am von Europa ausgegangenen Kolonialismus und der eng damit in Verbindung stehenden Sklaverei. Auch die vielen kriegerischen Auseinandersetzungen europäischer Mächte in den vergangenen Jahrhunderten waren begleitet von Kriegsverbrechen. Schließlich verstößt die vom Neoliberalismus geprägte Ausbeutung ehemaliger Kolonialgebiete bis heute an vielen Stellen gegen diese Werte.
Kolonialismus
Naturwissenschaftliche und technische Fortschritte ermöglichten es den Europäern ab dem 15. Jahrhundert, mit ihren Schiffen überseeische Gebiete zu erreichen und mit ihren Feuerwaffen zu erobern.
Die charakteristische Vorgehensweise, denen die Eroberer immer wieder folgten, war:
militärische Unterwerfung und Vernichtung der Infrastruktur
Kolonisierung mit Europäern
Unterwerfung, Umerziehung, Ausbeutung und teilweise sogar Vernichtung der indigenen Bevölkerungen
maximale Ausbeutung der Ressourcen mithilfe der zur Zwangsarbeit verpflichteten Einheimischen und importierten Sklaven
Vernichtung ursprünglicher Wirtschaftssysteme
Vernichtung ursprünglich vorhandener Ökosysteme
Aufbau einer supressiven Kolonialverwaltung
nach der „Freilassung“: Verhinderung einer wirklichen Unabhängigkeit durch die ehemaligen Kolonialherren in Zusammenarbeit mit korrupten „Ortskräften“.
Dabei wurde (und wird) gegen alle von der EU formulierten Grundwerte in eklatanter und grober Weise verstoßen. Diese Verstöße sind zahlreich belegt, dokumentiert und teilweise auch literarisch verarbeitet. Eine sehr gute Übersicht bietet das von Jean Ziegler 2008 verfasste Buch „Der Hass auf den Westen“.
Da die europäische Hauptstadt Brüssel in Belgien liegt, seien die Verbrechen in „Belgisch-Kongo“ als ein Beispiel heausgegriffen. In der von Otto von Bismarck geleiteten Kongokonferenz 1884/85 wurde der Kongo dem belgischen König Leopold II von den europäischen Nationen praktisch als Privatvermögen zugesprochen und euphemistisch als „Kongo-Freistaat“ bezeichnet. „Frei“ stand hier für großzügige Freihandelsbestimmungen, nach denen der Kongo für die europäische Wirtschaft geöffnet werden sollte. Dadurch konnte sich in den nächsten Jahren ein System unreglementierter Raubwirtschaft etablieren. Zunächst ging es vor allem um den Rohstoff Elfenbein, später dann im Zuge des Kautschukbooms um Naturkautschuk. Die indigene Bevölkerung wurde einem brutalen Kolonialsystem unterworfen. Körperliche Züchtigung, Verstümmelungen, Geiselnahmen und Morde waren die Methoden, mit denen versucht wurde, maximale Gewinne aus dem Land zu ziehen. Je nach Schätzungen sollen dabei bis zu 10 Millionen Einwohner ums Leben gekommen sein. Erst nachdem die Weltöffentlichkeit durch den Bericht des britischen Diplomaten Roger Casement über die Kongo-Gräuel aufgeschreckt worden war, wurde der „Freistaat“ dem Privatbesitz von König Leopold entzogen und direkt dem belgischen Staat unterstellt (vgl. Joseph Conrad: Herz der Finsternis, Mario Llosa Vargas: Der Traum des Kelten, Adam Hochschild: King Leopolds Ghost).
Doch auch unter der Regie des belgischen Staates ging die Ausbeutung des Kongo weiter. Neben Kautschuk, Palmöl und Kaffee spielten nun zunehmend auch Bodenschätze wie Kupfer, Blei, Zink und Diamanten eine wichtige Rolle.
Im Zweiten Weltkrieg wurde Uran aus dem Kongo für das amerikanische Atombombenprogramm genutzt, auch die Kupfer-, Palmöl-und Gummiindustrie erlebte einen Aufschwung. Nach wie vor wurde die afrikanische Bevölkerung nicht an einer Verwaltung beteiligt, was zu Unruhen führte. Zur Jahreswende 1941/42 wurden auf einem Fußballplatz in Elisabeth will mindestens 60 streikende Kongolesen erschossen.
In den Nachkriegsjahren verstärkte sich der Widerstand gegen die belgischen Kolonialregierung. Der belgische Staat versuchte, durch eine Reihe von Reformen diesen Widerstand zu begegnen. Gegen Ende der 1950er Jahre waren zwar zahlreiche Afrikaner an der Verwaltung beteiligt, aber bis zur Unabhängigkeit, bei der die Kolonie in Zaire umbenannt wurde, hatten Afrikaner kein volles Wahlrecht. Die mit Unruhen verbundenen Unabhängigkeitsbestrebungen wurden 1959 vom belgischen Staat zunächst noch hart bekämpft. Aber nachdem im Januar 1960 die französische Regierung das Nachbarland Kamerun in die Unabhängigkeit entlassen hatte, kündigte auch die belgische Regierung ihren Rückzug aus dem Kongo an.
Nach dem Ende des offiziellen Kolonialstatus am 30. Juni 1960 wurde Patrice Émery Lumumba zum Staatspräsidenten gewählt. Er versuchte, den Kongo auf friedliche Weise in eine Unabhängigkeit zu führen, in der die wirtschaftliche Nutzung von Bodenschhätzen und Rohstoffen verstärkt der kongolesischen Bevölkerung zu gute käme . Unter Mithilfe belgischer Behörden und westlicher Geheimdienste wurde er zunächst festgenommen und dann zusammen mit zwei seiner Gefolgsleute in Katanga gefoltert und ermordet. Die Leichen wurden zersägt und mit Batteriesäure aufgelöst, um Spuren zu verwischen. Sein korrupter Nachfolger Mobutu, der die weitere Ausbeutung des Kongo durch die alten und neuen Kolonialmächte zuließ und sich dabei enorm bereicherte, legte den Grundstein für die vielen Probleme, mit denen dieses an sich enorm reiche Land und seine Bevölkerung bis heute zu kämpfen haben.
Im September wird Lumumba zu Hausarrest verurteilt. Er versucht zu fliehen, doch Armeechef Mobutu lässt ihn von Soldaten verfolgen. Mit zwei seiner Mitstreiter wird Lumumba gefasst und in einer Kaserne bei Thysville im Westen des Landes interniert. Bald danach bringt man ihn auf Betreiben des belgischen Ministers d’Aspremont nach Katanga; er wird mehrere Stunden verhört und gefoltert. Im Beisein von vier belgischen Polizisten, Präsident Tschombé und zwei Ministern der abtrünnigen Provinz werden Lumumba und seine Mitstreiter erschossen. Um alle Spuren zu beseitigen, gräbt ein belgischer Polizeibeamter die Leichen kurze Zeit später wieder aus, zerstückelt sie mit einer Säge und löst sie in Säure auf. (aus: Der tödliche Mut des Patrice Lumumba von Alexander Behr, Deutschlandfunk Kultur ,13.01.2021)
Soviel zum Kongo. Dieses Land soll hier exemplarisch stehen für den „Werteexport“ aus Europa in europäische Kolonien. Auf einige weitere Beispiele sei nur kurz hingewiesen:
Die Gräueltaten der spanischen und portugiesischen Eroberer in Amerika wurden zum größten Teil mit Billigung der katholischen Kirche durchgeführt und teilweise damit entschuldigt oder begründet, dass man den einheimischen Menschen den Status des „Menschseins“ absprach (Disput von Valladolid, 1550/51). So wurden in den Silberminen des Cerro Rico bei Potosi im heutigen Bolivien in drei Jahrhunderten 40.000 t Silber abgebaut und dabei kamen fast 4 Millionen Indianer ums Leben (aus Ziegler 2008 nach Hamilton 1934). Die Arbeitsbedingungen der Mitayos, wie die Minenarbeiter genannt wurden, zu denen auch Frauen und Kinder gehörten, waren extrem grausam. Wer sich weigerte, die Leitern hinabzusteigen, wurde sofort erschossen und heraus durfte nur, wer ausreichend Mengen Silbererz mit sich brachte. Revolten und Aufstände wurden mit äußerster Brutalität niedergeschlagen.
Die Kolonisierung Nordamerikas war ebenfalls ein extrem gewalttätiger Akt, begleitet von Vertragsbrüchen, falschen Versprechungen und mörderischen Vertreibungen. In kurzer Zeit wurde ein Großteil der ursprünglich einheimischen Bevölkerung vernichtet. Der Ausrottungsprozess wurde durch eingeschleppte Krankheiten, zum Beispel der Malaria, erheblich beschleunigt (Diamond 1997). Die Zeit dieses vor allem im 18. und 19. Jahrhundert ablaufende Völkermords wird in dem bis heute recht beliebte Genre des Wildwestfilms mit seiner Schwarzweißmalerei und der durch lockere Colts durchgesetzten gerechten Sache nicht nur romantisch verzerrt und verharmlost, sondern völlig vernebelt und vertuscht
Eine besonders grausame Methode, „europäische Werte“ zu vermitteln, wurde vor allem unter englischer Kolonialherrschaft, zuerst in Australien und später auch in Kanada, betrieben. Um die Eingeborenen zu „zivilisieren“, wurden die Kinder ihren Eltern weggenommenn und in Erziehungsanstalten untergebracht. Über die grausamen Lebensbedingungen in diesen meist unter christlicher Oberhoheit stehenden Anstalten, die bis 1969 bestanden, hat man erst in jüngerer Zeit mehr erfahren (vgl. z. B. Read 1981, 2006).
Cootamundra Girls Home. Cootamundra Girls Home, established in 1911, was the first of the homes for Aboriginal children set up by the Aborigines Protection Board. The main aim of the Board was to ‚rescue‘ Aboriginal children from their families and assimilate them into the white community. Girls were the main target of the Board, especially so-called ‚half-caste‘ or ‚mixed blood‘ girls. The girls were trained as domestic servants and sent out to work for middle class white families.At Cootamundra, Aboriginal girls were instructed to ‚think white, look white, act white‘. This was part of the process to make the girls suitable wives for white men, in the hope that through interracial marriages, Aboriginal blood would be ‚bred out‘. They were taught to look down on their own people and to fear Aboriginal men.Girls in the home were not allowed to communicate with their families. They were often told that their parents were dead and even given forged death certificates. As a result, many of the girls in the home lost their families forever.Cootamundra Home was closed in 1968, the year before the Aboriginal Welfare Board (previously the Aborigines Protection Board) was abolished. https://web.archive.org/web/20081206184900/http://www.dreamtime.net.au/indigenous/family.cfm#bi
Russland errichtete zwar keine überseeischen Kolonien, kolonisierte dafür aber ganz Sibirien und den nordpazifischen Raum bis Alaska. Auch dabei wurde und wird die indigene Bevölkerung unterworfen, unterdrückt und ausgebeutet (Zeugnisse dafür finden sich zum Beispiel bei Georg Wilhelm Steller und Adelbert von Chamisso).
Sklaverei
Menschen im Besitz anderer Menschen, also Sklaven, gab es schon im alten Ägypten. In ganz großen Maßstab wurde Sklaverei und Sklavenhandel aber zwischen Europa, Afrika und der „Neuen Welt“ im 16. Jahrhundert begonnen und erst im 19. Jahrhundert gegen große Widerstände beendet. Dabei waren auch afrikanische Herrscher und Zulieferer beteiligt, die entscheidende Triebkraft für die riesengroße Menge an Sklaven, die in die neue Welt verschifft wurden, ging aber von spanischen, französischen und englischen Sklavenhändlern aus.. Dabei spielte der Dreieckshandel – Waffen und Söldner von Europa nach Afrika, Sklaven von Afrika nach Amerika, Zucker von Amerika nach Europa – eine wichtige Rolle. Von den etwa 50 Millionen von Afrika nach Amerika gebrachten Sklaven starben 10-20 % schon bei der Überfahrt.
Wichtigster Teil der Unabhängigkeitserklärung der Vereinigten Staaten von Amerika von 1776 ist die darin enthaltene Formulierung der Menschenrechte. Der Hauptautor Thomas Jefferson besaß bei seinem Tode 1826 200 Sklaven!
Europäische Kriegsverbrechen und Genozide
Viele innereuropäischen Kriege wurden mit großer Grausamkeit geführt. Besonders hervorzuheben ist der 30-jährige Krieg, aber auch in den 100 Jahre andauernden kriegerischen Auseinandersetzungen zwischen England und Frankreich im 14. und 15. Jahrhundert kam es zu vielen Kriegsverbrechen. Den bisherigen Gipfel der Grausamkeiten stellt der während des Zweiten Weltkriegs unter den deutschen Nationalsozialisten geplante und durchgeführte Genozid an Juden und weiteren „nicht arischen“ Menschen dar.
„Die Schuld des Nordens“
Sind dies alles Ereignisse der Vergangenheit? Hat – zumindest mit dem Vertrag von Lissabon von 2009 – eine neue humanistische Ära in Europa angefangen?
Ausbeutung, Armut, Hunger und Not haben auf der Welt in den letzten Jahrzehnten nicht abgenommen. Ob man die Arbeiterinnen in den Textilfabriken Bangladeschs, die Kinderarbeit in Kakaoplantagen Afrikas und Südamerikas, die rechtswidrigen, Umwelt und Menschen zerstörenden Formen der Ölförderung im Nigerdelta oder die unmenschlichen Bedingungen des Coltan-Abbaus in der Demokratischen Republik Kongo als Beispiel nimmt, immer sind amerikanische und europäische Firmen Nutznießer und Beteiligte und widersprechen mit ihren Handlungen den formulierten EU-Grundrechten. Es gibt zwar sehr viele Bemühungen, die Situation zu verbessern – man denke etwa an das „Lieferkettengesetz“ – aber es ist ausgesprochen schwierig, humanistische Ziele und Menschenrechte gegen wirtschaftliche Interessen durchzusetzen. In dieser Beziehung hat sich seit dem 19. Jahrhundert nicht sehr viel geändert. Ein Unterschied ist vielleicht, dass heute große, multinationale Konzerne – zumindest in der westlichen Welt – mehr Einfluss haben als Regierungen.
In jedem Fall sollte man heute und in Zukunft vermeiden, die Verbreitung „europäischer Werte“ als wichtiges politisches Ziel zu verkünden. Alle Nicht-Europäer müssen diese Bezeichnung als Hohn empfinden.
Humanistische Werte und Menschenrechte, um die es gehen sollte, sind alles andere als „europäisch“. Die mildeste Verurteilung dieser Bezeichnung ist Scheinheiligkeit aber eher noch handelt es sich wohl um bewusste Irreführung.
Quellen
Behr, A. (2021): Der tödliche Mut des Patrice Lumumba. Deutschlandfunk Kultur,13.01.2021
Bitterli, U. (1976): Die „Wilden“ und die „Zivilisierten“. München: C. H. Beck
Chamisso, A. v. (2001): Reise um die Welt. Aufbau Taschenbuch
Conrad, J. (2005): Herz der Finsternis. München: dtv; Erstveröff. „Heart of Darkness“, 1899
Crosby, A. W. (1991): Die Früchte des weißen Mannes: ökologischer Imperialismus 900-1900. Frankfurt a. M. : Campus
Diamond, J. (2006): Arm und Reich.7. A., Frankfurt: Fischer; Erstveröff. „Guns, Germs and Steel“ 1996
Fanon, F. (1969): Die Verdammten dieser Erde. Rororo-Taschenbuchausgabe; Erstveröff. „Les damnés de la terre“,1961
Hochschild, A. (1999): King Leopolds Ghost. Mariner Books, Reprint Edition
Mann, C. C. (2013): Kolumbus‘ Erbe. Reinbek: Rowohlt
Probst, W. (2015): Der Palme luft’ge Krone – mit Chamisso auf Weltreise, 2. A..Ochsenhausen: Angele-Verlag
Read, P. (1981, 2006): The stolen generation. The removal of Aboriginal children in New South Wales 1863 – 1969. New South Wales Departmet of Aboriginal Affairs. https://www.aboriginalaffairs.nsw.gov.au
Sabet, H. (1992): Die Schuld des Nordens.2. A., Frankfurt am M.: Horizonte-Verlag
Steller, G. W. (1986): Von Sibirien nach Amerika. Hrsg. V. Matthies. Stuttgart/Wien: Thienemann
Vargas Llosa, M. (2012): Der Traum des Kelten. Hamburg: Suhrkamp; Erstveröff. „El seño del celta“, 2010
Ziegler, J. (2008): Der Hass auf den Westen, 2. A.. München. Bertelsmann
Zusatzinformationen zum Basisartikel von UB 475 „Wasserhaushalt der Pflanzen“
Titelbild: Sonnenblume, Netztracheide längs, gekreuzte Polfilter (Foto W. Probst)
Die ursprünglichen Wasserleitungszellen der Pflanzen waren lang gestreckt, verhältnismäßig dünn und mit sehr schräg stehenden Querwänden versehen. Die stammesgeschichtliche Entwicklung brachte vor allem eine Erweiterung des Zellvolumens, oft bei gleichzeitiger Verkürzung der Zellen. Entscheidend für die phylogenetische Entwicklung der Xylemelemente war die Differenzierung der sekundären Wandstrukturen.
Während für die Wasserleitungselemente der Moose schon ein teilweiser Abbau der Querwände aber noch keine Lignineinlagerung und keine sekundären Wandversteifungen nachgewiesen sind, sind solche Tracheiden und Tracheen typisch für die Gefäßpflanzen (Tracheophyta).
Ring- und Schraubentracheiden
Das Xylem der ausgestorbenen Urfarne (Psilophytopsida) bestand ausschließlich aus Tracheiden mit ring- oder schraubenförmigen Wandverdickungen. Da soche Wandversteifungen auch bei den heutigen Gefäßpflanzen in der Ontogenie meist zuerst angelegt werden, gelten sie als ursprünglich.
Treppentracheiden
Werden horizontal liegende Versteifungsringe durch vertikale Stege verbunden, so entstehen großlumigen Tüpfel. Bei der Treppentülpfelung liegen seitlich benachbarte Tüpfel stets auf gleicher Höhe. Je nach Anzahl der Vertikalbrücken unterscheidet man einreihige und mehreihige Treppentüpfel.
Netztracheiden
Netztüpfel kann man sich durch Anastomosen zwischen mehreren zum Teil gegenläufigen Schraubenversteifungen entstanden denken. Seitlich benachbarte Tüpfel scheinen hier in der Höhe gegeneinander versetzt. Eine unregelmäßige Anordnung der einzelnen Tüpfel gilt als ursprünglich, eine regelmäßige Anordnung in Längsreihen als abgeleitet.
Phylogenie und Ontogenie der Wasserleitunngsbahnen der Pflanzen
Behöfung der Tüpfel
Bereits Ringtracheiden zeigen eine Tendenz zur „Behöfung“: Die Verstärkungsleiste besteht aus einem Leistenfuß, dem der breitere Leistenkörper flanschartig aufgesetzt ist. Bei Treppen- und Netztüpfeln führt die überragende Sekundärwand dazu, dass ein Binnenraum über der Primärwand (Mittellamellen, Schließhaut) gebildet wird, der nur noch über relativ enge Poren mit dem Zellvolumen in Verbindung steht.
Mögliche pylogenetische Entwicklung von Hoftüpfeln aus Teppentracheiden (Beide Abbildungen aus Probst, W. (1987): Biologie der Moos- und Farnpflanzen. 2. A., Heidelberg/Wiesbaden: Quelle und Meyer)
Tracheen und Tracheiden
Während man lang gestreckte Wasserleitungsgefäße mit Querwänden als Tracheiden bezeichnet, werden Gefäße mit aufgelösten Querwänden Tracheen genannt. Stammesgeschichte stellt man sich eine Entwicklung über Treppentracheiden vor, bei denen zunächst die Schließhaut, dann auch die Leitersprossen aufgelöst wurden.
Programm (Stand 12.04.2022), alle Exkursionen finden am Samstag statt, bitte an wetterangepasste Kleidung und stabile wasserfeste Schuhe, Sonnenhut und Sitzkissen denken. Möglichst Fernglas, Botanikerlupe, Kamera und Schreibzeug mitbringen. Exkursions- und Arbeitsmaterialien werden von den Dozenten mitgeführt oder vor den Exkursionen per E-Mail mitgeteilt. Vesperpause vor Ort während der Exkursion.
Wälder, Wiesen, Weiher am Stillen Bach und Rössler Weiher, Frühjahrsblüher und Gehölze im Wald, Streuobstwiesen, Singvögel, Wasservögel, Vogelzug, Amphibien, Arthropoden und Bodenfauna
Referent: Bertrand Schmidt
Achtung!Terminänderug wegen Regenwetter:
14.05.2022, Wanderparkplatz bei Appenweiler K7731, 13.30-17.30 Uhr
Lebensform Baum, ökologische Ansprüche von Waldbäumen und Waldkräutern, Gräben im Wald, Tierspuren
Nass- und Streuwiesen im Naturschutzgebiet Lochmoos und Altdorfer Wald, Schutzgebietsmanage-ment, Biberrevier mit Biberburg, Pflanzen, Amphibien, Falter, Ökologie der Feuchtgebiete
Wanderparkplatz Steinach, Riedweg (Gde. Bad Waldsee)
Der Treffpunkt Parkplatz Nessenreben, Weingarten Schwimmbad (ohne Plan) ist über den Routenplaner leicht zu finden. Er liegt in 1 km Entfernung von der PH-Weingarten.
Treffpunkte Exkursionen mit Herrn Probst
Adelsreuter-Weißenauer Wald, Waldparkplatz an der K7731 östlich Appenweiler
Fahrt von Ravensburg auf der B 33 Richtung Meersburg bis Dürnast, dort links abbiegen nach Taldorf und weiter bis Appenweiler, dort links abbiegen Richtung Brochenzell, nach etw 300 m ist links ein kleiner Parkplatz am Waldrand
Oberteuringen, Ecke Jahnweg/Franz-Roth-Platz
Fahrt von Ravensburg auf der B 33 Richtung Meersburg bis Oberteuringen/Neuhaus, dort links ab auf der Teuringer Straße,
… die hinter dem Ortsschild in die Augustin Bea-Straße übergeht. Der Franz Roth-Platz liegt links.
Informationen zu übergeordneten Inhalten und Zielen
Die große Bedeutung, die der biologischen Vielfalt für eine nachhaltige Entwicklung unseres Bioplaneten gegeben wird, steht in keinem Verhältnis zum Bildungsstand der Bevölkerung. Artenkenntnisse und grundlegende Kenntnisse heimischer Ökosysteme sind deshalb ein wichtiger Teil der Allgemeinbildung, die durch Biologieunterricht vermittelt werden sollte. Und sie müssen deshalb auch ein wichtiger Bestandteil der Lehrer*innenausbildung sein. Es ist naheliegend und durch viele empirische Untersuchungen belegt, dass sich diese Kenntnisse am besten im Freien, also auf Exkursionen, erreichen lassen. Dabei geht es um alle Aspekte der biologischen Vielfalt.
Ein zentrales Ziel ist die Verbesserung der Formen- und Artenkenntnis. Dieses Ziel lässt sich natürlich durch sechs Exkursionen alleine nicht erreichen. Sie können nur Anstoß sein für eigene Aktivitäten. Ein erster Schritt ist die gründliche Nachbearbeitung anhand von Artenlisten, die für jede Exkursion aufgestellt werden. Kennt man einen Artnamen, so lassen sich mit dessen Hilfe über das Internet sehr viele Informationen über die Biologie und die ökologischen Ansprüche dieser Art gewinnen. Die Standortansprüche von Pflanzen werden sehr gut durch die auf empirische Grundlagen beruhenden Zeigerwerte (nach Ellenberg) wiedergegeben, die ebenfalls über das Internet zugänglich sind.
Diese ökologischen Ansprüche stehen in enger Beziehung zu Lebensraum und Lebensgemeinschaft, also zum Ökosystem, in das die jeweiligen Arten eingebunden sind. Unsere Exkursionen sollen deshalb auch einen Einblick in die Vielfalt der regionalen Ökosysteme vermitteln. Dabei ergeben sich enge Beziehungen zur Landschaftsgeschichte Oberschwabens, für die vor allem die letzte Kaltzeit (Würm) entscheidende Voraussetzungen lieferte. Die weitere nacheiszeitliche Entwicklung war ebenso geprägt von natürlichen Vorgängen wie von menschlichen Einflüssen. Diese menschlichen Einflüsse haben mit zunehmender Bevölkerungsdichte und vor allem mit Veränderungen der Siedlungsstruktur und der Landbewirtschaftung immer stärkerem Einfluss gewonnen und sind mittlerweile eine Bedrohung für die biologische Vielfalt geworden. Dies gilt nicht nur lokal, sondern global, was besonders an der treibhausgasbedingten Klimaerwärmung deutlich wird. Naturschutz bedeutet deshalb nicht nur den Schutz einzelner Arten, sondern vor allem den Schutz von Funktionsabläufen. Naturschutzmaßnahmen sollen helfen, regional und überregional eine für die Menschheit nachhaltige Entwicklung zu gewährleisten.
Der dritte Teilbereich der biologischen Vielfalt betrifft die genetische Vielfalt, die sich zum Beispiel in der Individualität aller Lebewesen ausdrückt. Diese genetische Vielfalt wird – wie die Artenvielfalt – durch Nivellierung bestimmter Umweltfaktoren gefährdet. Wichtigster Punkt ist hier das hohe Nährmineral-Angebot aller terrestrischen und aquatischen Lebensräume. Es gibt kaum noch oligotrophe Gewässer, Magerrasen oder Hochmoore.
Methoden
Selbständiges bearbeiten von Aufgaben, Gruppenarbeit, Naturerlebnisse; für jede Exkursion erstellen einer Artenliste
Studienleistung
Anfertigen eines Exkursionsprotokolls, auch in Form eines Portfolios, in dem zu jeder Exkursion mindestens zwei Objekte, Themen oder Aktivitäten etwas ausführlicher ausgearbeitet werden.
Bei der Erstellung eines Portfolios sollten folgende Aspekte beachtet werden: Verbindung des Lernprozesses und des Lernproduktes, Handlungsorientierung , selbstreflexivität des Lernens, Kompetenz- statt Defizitorientierung
Die Protokolle/Portfolios sollen bis Ende Juli – am besten in elektronischer Form – abgegeben werden.
Wetter
Für mich ist Regen sehr schlecht, ich muss mir deshalb die Möglichkeit offenhalten, bei Regenwetter Exkursionstermine zu verschieben. Das werde ich über E-Mail mitteilen.
Kommentare und Fragen – während den Exkursionen und danach – sind erwünscht!
14. und 16. Mai 2022 – Im Frühlingswald bei Appenweiler
Der Adelsreuter-Weißenauer Wald
Die Artenzusammensetzung dieses Waldes ist sehr vielfältig. Das lichte Hellgrün frisch ausgetriebener Laubbäume mischt sich mit verschiedenen Nadelbäumen und vielen verschiedenen Straucharten an Waldrändern und Wegrändern. Vegetationskundlich gesehen handelt es sich um einen Buchen-Tannenwald, dessen Artenzusammensetzung aber durch forstliche Maßnahmen ergänzt bzw. verändert wurde.
Größere zusammenhängende Waldgebiete sind im heutigen Oberschwaben ziemlich selten. Im Laufe der mehrtausendjährigen Siedlungsgeschichte ist die typische kleinräumige Landschaft aus Feldern – heute vielfach Obstplantagen -, Wiesen und kleinen Waldstücken entstanden. Das größte Waldgebiet, der Altdorfer Wald etwa zwischen Vogt und Wolpertswende gelegen, hat immerhin eine Längsausdehnung von ca. 17 km. Dagegen ist das Waldgebiet , das sich am westlichen Rand des Schussenbeckens etwa von Ravensburg bis Meckenbeuren erstreckt, mit knapp 8 km deutlich kleiner. Die geplante Umgehungsstraße für Meckenbeuren könnte es noch weiter verkleinern.
Trotzdem kann man in diesem Wald stundenlang wandern. Mehrere Bäche – zum Beispiel der von Appenweiler kommende Mühlbach – entwässern das Gebiet zur Schussen hin. Sie haben sich zum Teil ziemlich tief in die Jungmöräne eingeschnitten.
Exkursionsweg und wichtige Stationen
Ausschnitt aus der Topographischen Karte 1 : 25.000, Blatt 8223 Ravensburg
(1) Bäume blind ertasten und wiederfinden
Material: Schlafbrillen, Leintuch, Namensschilder für Baumarten
Schärfung des Tastsinns bei Ausschließen des optischen Sinns; erhöhte Geräuschempfindlichkeit
Auswertung: Jede*r bringt von seinem Baum etwas Charakteristisches >> Objekt auf Laken auslegen, Artenunterschiede, Individualität der Baumarten: Beschaffenheit der Borke, besondere Spuren; z.B. Astlöcher, Verletzungen, Tierspuren.
Auf dem rötlich braun gefärbten, ziemlich trüben Wasser tummeln sich zahlreiche Wasserläufer. Immer wieder kommen dunkle Tiere an die Oberfläche, die wir aber nicht fangen können (vermutlich Ruderwanzen). Die Wasserfarbe kommt durch oxidierte Eisenionen zustande.
Um den ganzen Grabenbereich wachsen dichte Bestände der Schlank-Segge (Carex acuta) – Sauergras mit dreikantigen Stängel und gleichzeitig gestellten Blättern, Vertreter der Verschiedenenährigen Seggen, bei denen männliche und weibliche Blüten in unterschiedlichen Ähren angeordnet sind. Bei den Gleichährigen Seggen enthalten alle Ähren männliche und weibliche Blüten. Wir registrieren: Winkel-Segge (Carex remota) und Zittergras-Segge (Carex brizoides).
Pflanzen mit besonderem Duft:
Ross-Minze (Mentha longifolia), einer Stammart der Pfeffer-Minze (Mentha x piperita)
Mentha x piperita entstand aus Mentha x spicata x Mentha aquatca, Mentha x spicata aus Mentha longifolia x Mentha rotundifolia
Sumpf-Mädesüß (Filipendula ulmaria): vor allem die unterirdischen Pflanzenteile riechen stark nach Methylsalicylsäure (=Salicylsäuremethylester oder Methylsalicylat), wegen seiner desinfizierenden Wirkung als Zusatzstoff in Zahnpasta und Kaugummis eingesetzt.
(3) Bäume zählen
Ziel: quantitative Abschätzung der Baumarten-Zusammensetzung eines Waldes
Wie könnte man vorgehen? Eine Möglichkeit wäre die Auszählung mehrerer Probequadrate, Mittelwertbildung und eure gesamte Waldfläche. Wir wählen die Transitmethode.
Material: Leintuch; mindestens 30 m lange Wäscheseile
Man kann darauf verzichten und sich stattdessen an die Regel halten, in jeder Zweiergruppe Blätter von 10 Baumindividuen einzusammeln, die etwa in einer Linie senkrecht zum Weg stehen.
Ergebnis unerer Zählung am 16.5.2022 (3 Gruppen)
Auswertung: Die Blätter (oder andere Objekte einer Baumart) werden zunächst am unteren Ende des Leintuchs nach Arten sortiert, dann werden die Blatthaufen der Größe nach geordnet und dann werden die Blätter zu Säulendiagramm angeordnet. Dabei muss darauf geachtet werden, dass für jedes Blatt/Objekt unabhängig von seiner Größe gleich viel Platz verwendet wird. So erhält man ein Säulendiagramm der Artenhäufigkeit.
(4) Berechnung der Kohlenstoffspeicherung von Bäumen
Ziel: quantitative Abschätzung der Kohlenstoffspeicherung durch Laubbäume
Material: Arbeitsblatt, Zollstock, 2-3 m lange Schnur,
Wie geht man vor? Besprechung auf der Grundlage des Arbeitsblattes
Auswertung: Wann sind Wälder Kohlenstoffsenken, wann Kohlenstoffquellen? Diskussionen zur Waldbewirtschaftung; eventuell Abschätzung: wie viel CO2 wird von einem Pkw bei der Fahrt von Weingarten nach Appenweiler freigesetzt.
(5) Lebensräume der Gelbbauchunke
In ausgedehnten Pfützen auf dem Weg und im angrenzenden Graben können wir Gelbbauchunken beobachten. Es sind sehr kleine Tiere, vermutlich vom letzten Jahr. Unken werden erst im zweiten Jahr geschlechtsreif gereichen dann ihre Größe von bis etwa 5 cm. Kaulquappen konnten wir nicht sehen, am Samstag beobachteten wir auch einige größere Tiere.
(6) Wachstum und Alter von Bäumen
Ziel: Altersbestimmung von Bäumen durch Auszählen der Jahresringe
Material: Stecknadeln, Papierfähnchen zur Beschriftung
Auswertung: Aus der Dicke der Jahresringe kann man Aussagen über die Wachstumsbedingungen machen; Jahresringechronologie; zum Auszählen der Jahresringe verwendet man Stecknadeln, die man alle fünf oder zehn Ringe einsteckt; man kann besondere Daten mit Fähnchen versehen (zum Beispiel Geburtsjahre)
Rinde aus Borke und Bast, Kambium, Holz; Kambium; Periderm: v.i.n.a. Phelloderm,Phellogen (Korkkambium),Phellem, Lentizellen.
Altersbestimmung ens Lärchenstammes: Alle 5 Jahre wird eine blaue Stecknadel gesteckt, die gelben Nadenmmarkieren Geburtsjahre von teilnehmerinen (Foto Dominique Mattivi)
(7) Ess-Kastanien und Rosskastanien
An der Wegeinmündung stehen sich eine Rosskastanie und eine Ess-Kastanie gegenüber. Warum die unterschiedliche Schreibweise? Die beiden „Kastanien“ sind nicht mehr miteinander verwandt. Die Rosskastanie, genauer die Art Gewöhnliche Rosskastanie (Aesculus hippocastanum) gehört zu den Seifenbaumgewächsen, „Rosskastanie“ bezeichnet also eine Gattung. Die Ess-Kastanie (Castanea sativa) auch auch Echte Kastanie genannt, bezeichnen eine Art aus der Gattung Kastanie . „Ess-“ bzw. „Echte“ ist der Artzusatz oder das Epitheton. Wenn man auf diesen Unterschied zwischen Art- und Gattungsbezeichnung auch bei der Schreibweise der deutschen Namen achtet, geben Sie den Sachverhalt so exakt wieder wie die wissenschaftlichen Namen. In wissenschaftlichen Bestimmungsbüchern wird dies deshalb normalerweise so gehandhabt.
(8) Mein Kraut in der Suppe
Jahrhunderttausende lebten die Menschen und ihre Vorfahren als Jäger und Sammler, vermutlich meist mehr als Sammler. Sie ernährten sich von Wildpflanzen, ihren Früchten, Speicherorganen, aber auch Blättern und Blüten. Das Sammeln ist also die ursprünglichste Form der Nahrungsbeschaffung. Erst vor 10-15.000 Jahren haben die ersten Menschen damit begonnen, Ackerbau zu betreiben und Haustiere zu züchten. Man kann deshalb davon ausgehen, dass sich die lange Praxis des Sammelns von Nahrungsmitteln auch in den Genen des Menschen niedergeschlagen hat, dass es eine genetische Disposition zur Sammelleidenschaft gibt. Besonders deutlich wird das vielleicht heute noch beim Pilzesammeln.
Aber auch ein selbst gesammeltes Kraut zu essen kommt einem sehr ursprünglichen Bedürfnis entgegen. Außerdem verbindet man mit Kräutern meist, dass diese etwas Gutes tun, der Gesundheit, dem Aussehen oder dem Wohlbefinden förderlich sind. Diese überragend wichtige Rolle entspricht der traditionellen Bedeutung der Kräuter in unserer Gesellschaft, die sie allerdings in den letzten 100 Jahren eingebüßt haben. Gegenwärtig erfährt das Wildkräuter sammeln. Ihre Verwendung in der Küche allerdings eine Renaissance. Das kann man an der sehr großen Zahl entsprechender Bücher mit Sammelanleitungen und Rezepten ableiten. Trotzdem kennen die meisten Leute Kräuter nur von der Kräuterecke im Supermarkt, allenfalls noch vom eigenen Garten, kaum aus der freien Natur. Die uralte Tradition der Pflanzenkenntnis – Erfahrungen, die über Jahrtausende weitergegeben wurden – sind weit gehend verloren gegangen. Wenn wir Kräuter sammeln, um sie nachher zu essen, dann treten wir in eine uralte Tradition ein. Kindern und Jugendlichen kann diese Übung den Zugang zur Pflanzenvielfalt erleichtern.
Vorgehensweise
Sammeln („Sammle fünf Kräuter, die du gerne essen würdest“)
Bestimmen (Die gesammelten Kräuter werden nach Arten sortiert und bestimmt. Die Bestimmungsergebnisse sollten durch einen Experten abgesichert werden)
Auswählen (Einige essbare Arten werden ausgewählt; Informationen über die Verwendbarkeit man zum Beispiel im Internet: Eingabe: Pflanzenname und „essbar“)
Ergänzung für den Kochtopf (Von den ausgewählten Arten wird so viel gesammelt, wie für die Zubereitung einer entsprechenden Mahlzeit benötigt wird)
Aus hygienischen Gründen ist es ratsam, im Wald oder im Gelände gesammelte Kräuter nicht roh zu genießen, also zum Beispiel in Salaten, Kräuterquark oder Smoothies. Empfehlenswert sind Kräutersuppen oder Gemüse, bei denen das Sammelgut nach dem Waschen auch noch erhitzt werden muss.
Tierspuren
Minen
„Minen [von franz. mine = Bergwerk], Hyponomien, Nomien, durch Fraß (Minierfraß) von meist Larven der Insekten (Minierer) in parenchymführenden Pflanzenteilen (Grundgewebe) entstandene Hohlräume, deren Form, gelegentlich auch die Art und Weise der Kotablagerungen, oft schon die Identifikation der Erzeuger (Zweiflügler, Klein-Schmetterlinge, Käfer, Blattwespen) ermöglicht“ (Definition aus Spektrum Lexiokon der Biologie).
Gänge von Larven der Geißblatt-Minierfliege (Phytomyza xylostei) auf dem Blatt der Roten Heckenkirsche (Lonicera xylosteum); eine solche Mine haben wir auf der Exkursion entdeckt. (Foto W. Probst)
Pflanzengallen
Pflanzengallen sind besondere, oft sehr differenzierte Strukturen, die von Pflanzen aufgrund von Reizen gebildet werden, die vonTieren oder Pilzen ausgehen. Die Bildungen dienen diesen Parasiten als Nahrung, zum Beispiel den Larven der Buchengallmücke (Mikiola fagi), die ihre Eier in Buchensblätter legt. Den Pflanzen nützen die Gallbildungen nichts, weshalb der Philosoph Erich Becher auh von„fremddienlicher Zweckmäßigkeit“ gesprochen hat.
Außer den Zitzengallen der Buchengallmücke haben wir auch noch Gallen an den Blättern des Giersch beobachtet, die von Blattflöhen (Trioza flavipennis) verursacht werden.
Gallen des Blattflohs Trioza flavipennis am Giersch (Foto W. Probst)
Blattfraß
Die häufigsten Tierspuren an Pflanzen, insbesondere an Blättern, sind Fraßspuren. Im Mai kann man fast an jedem Strauch des Gewöhnlichen Schneeballs (Viburnum opulus) die Fraßspuren des Schneeballblattkäfers (Pyrrhalta viburni) beobachten. Und wenn man genauer hinschaut– zum Beispiel mit der Botanikerlupe –, kann man auch die kleinen, schwarzgrauen Käferlarven erkennen.
Zusätzliche Infos und Materialien zu den Exkursionsthemen
Die Lebensform Baum
Wachstum
Zuwachsschichten eines Baumes
Alle Pflanzen wachsen, solange sie leben. Dieses Wachstum geht in jedem Fall von den Bildungsgeweben (Meristemen) an den Spitzen der Wurzeln und Sprossachsen aus. Bei vielen Pflanzen gibt es außerdem ein sogenanntes sekundäres Dickenwachstum, für das ein ringförmiges Bildungsgewebe verantwortlich ist, das Kambium. In den Bildungsgeweben finden ständig Zellteilungen statt. Durch starkes Streckungswachstum der jungen Zellen kommt dann der eigentliche Wachstumsschub zustande. Während dieses Streckungswachstums differenzieren sich die Zellen in unterschiedliche Gewebe. Während das Längenwachstum eines Baumes und eines Zweiges auf die Spitzenregion beschränkt ist, sind alle Achsenorgane in ihrer gesamten Länge zu Dickenwachstum in der Lage. Diese kontinuierliche Verdickung führt dazu, dasProbst)s ein Stamm einer zweikeimblättrigen Pflanze – dies gilt ebenso für die Nacktsamer – in seinen ältesten, also untersten Teilen, am dicksten ist und nach oben – entsprechend der Zeit, die das Kambium tätig war – immer dünner wird. Durch die Aktivität von Gipfelmeristem und Kambium wird jedes Jahr ein Zuwachskegel gebildet.
Da während der Wintermonate ein Wachstumsstopp eintritt und auch in den übrigen Jahreszeiten der Zuwachs nicht ganz gleichmäßig verläuft, kann man im Querschnitt jährliche Zuwachsringe beobachten. Mit diesen Jahresringen lässt sich das Alter eines Stammes oder Astes bestimmen. Aber je nach Umweltbedingungen fallen diese Zuwachsringe auch unterschiedlich dick aus. Sie sagen deshalb auch etwas die Lebensgeschichte des Baumes, Klima, Krankheiten Konkurrenz aus.
Das doppelte Missverständnis von E. O. Plauen
Borkenbildung
Borkenbildung
Nach innen werden von dem Kambiumring wasserleitende Elemente abgegeben – das sogenannte Holz oder Xylem (von gr. xylon = Holz) – nach außen das Assimilate Leitende Phloem (von gr. phlóos = Bast, Rinde).
Mit zunehmendem Dickenwachstum wird die primäre Rinde des jungen Stammes gesprengt. Es muss deshalb ein sekundäres Abschlussgewebe entwickelt werden. Dies geschieht über ein weiteres neu entstandenes Bildungsgewebe, das sogenannte Korkkambrium. Da sich aus bereits differenzierten Rindenzellen entwickelt, spricht man auch von einem sekundären Bildungsgewebe. Dieses Phellogen oder Korkkambium gibt nach außen meist verkorktes Phellem und nach innen Phelloderm ab. Die drei Gewebe zusammen werden als Periderm bezeichnet. Dieses Gewebe kann langlebig sein – zum Beispiel bei der Rot-Buche – oder schon nach einigen Jahren absterben und dann in tiefer liegenden Rindenschichten durch ein neues Periderm ersetzt werden. Dieses Folgeperiderm kann ringförmig oder schuppenförmig angeordnet sein. Durch die undurchlässigen Phellemschichten sterben die außerhalb liegenden Gewebe ab und werden bei weiterem Dickenwachstum meistens rissig (Borkenbildung).
Da diese Borkenbildung sehr unterschiedlich ablaufen kann, unterscheiden sich die Oberflächen von Stämmen, Ästen und Zweigen der Bäume stark. Sie sind ein wichtiges Merkmal zur Unterscheidung von Arten, aber sie unterscheiden sich auch von Individuum zu Individuum. Den die Veränderungen der äußeren Rinde sind abhängig von der Lebensgeschichte und den Umweltbedingungen des jeweiligen Baumes. Eine große Rolle spielt der Standort (Wald oder freistehend, Hang oder Ebene), aber auch der Einfluss von Krankheiten und anderen tierischen oder pflanzlichen Lebewesen und mechanischen Verletzungen.
Die Rinde umgibt den ganzen Baum oder Strauch und schützt ihn vor physikalischen oder biologischen Umwelteinflüssen. Schnee habe Wärmefrost UV-Strahlen oder schädliche Chemikalien werden durch die Rinde vom empfindlichen Kambium ebenso abgehalten, wie Insekten, Vögel, Säugetiere, Pilze und Bakterien. Deshalb bedeuten Verletzungen der Rinde für Bäume eine ähnliche Gefahr wie Hautverletzungen bei Menschen.
Eine weitere wichtige Funktion ist die Speicherung von gefährlichen Abfallstoffen in den abgestorbenen äußeren Bereichen der Rinde, der Borke. Die Gerbstoffe, Schleime und Harze dienen gleichzeitig als Abwehrstoffe gegen Schädlinge.
Kohlenstoffspeicherung
Es ist unbestritten, dass Wälder den größten Teil der in terrestrischer Biomasse enthaltenen rund 2000 Gt (Gitatonnen) Kohlenstoff speichern. Dafür ist vor allem das Holz verantwortlich. Im Laufe der Menschheitsgeschichte musste ein großer Teil der Wälder der landwirtschaftlichen Nutzung weichen. Dieser Prozess hat in den alten Kulturregionen Naher Osten, Indien, China und dem Mittelmeerraum schon vor mehreren Jahrtausenden begonnen. Dann setzte er sich im nördlichen Europa und in den Kolonialgebieten fort und erfuhr durch die Industrialisierung eine weitere Beschleunigung. Der Erhalt von Waldökosystemen ist deshalb auch im Sinne des Klimaschutzes ein wichtiges Ziel.
Für die Kohlenstoffbilanz ist aber auch wichtig, ob die natürlichen Speicher schon voll sind bzw. ihre maximal mögliche Größe erreicht haben, oder ob sie noch mehr Kohlenstoff aufnehmen können. Zum zweiten ist von Bedeutung, inwieweit auch das geerntete Holz als Kohlenstoffspeicher erhalten bleibt.
Durch Naturschutzmaßnahmen und Aufforstungsprogramme gibt es verschiedene Möglichkeiten, die natürlichen Speicher zu vergrößern:
Erhöhung der Speicherkapazität durch Waldschutz
Erhöhung der Biomasse in Wirtschaftswäldern durch forstwirtschaftliche Maßnahmen
Aufforstung bisher waldfreier Gebiete
Zum zweiten Punkt kann die Verwendung von Holz als langlebiger Rohstoff beitragen. Dies gilt am ehesten für Bauholz (ca. 100 Jahre und mehr), in geringerem Ausmaß für Holz, das für Möbel und andere Gebrauchsgegenstände genutzt wird (ca.20 Jahre, beide Werte nach Schönauer 2011, Bundestagsvorlage), noch weniger für Holz, das für die Papierherstellung genutzt wird. Da derzeit allerdings in Deutschland fast 70 % des Papiers recycelt werden (Papierkompass 2013), ergibt sich auch hier noch ein gewisser Speicherwert. Dabei muss man allerdings den hohen Energiebedarf der Papierfabriken berücksichtigen. Selbst für Brennholz lässt sich ein positiver Wert errechnen, wenn man davon ausgeht, dass es als Ersatz für fossile Brennstoffe genutzt wird. Die Kohlenstoffmenge, die ein Baum speichert, lässt sich durch ein relativ einfaches Verfahren abschätzen.
Abschätzung der Kohenstoffspeicherung in Bäumen
Beispiele für essbare Wildkräuter
Art
Essbare Pflanzenteile
Verwendung
Acker-Gänsedistel
weiche junge Blätter (bitterer Geschmack)
Salate, Gemüse, Suppen
Acker-Kratzdistel
junge Sprosse und Blütenkopfstiele
Gemüse, Suppen
Bachbungen-Ehrenpreis
junge Blätter und Sprosse vor der Blüte
Salatwürze, Suppen
Barbarakraut
junge Blätter
Salate, Gemüse, Suppen, (senfartiger Geschmack)
Blutweiderich
junge Blätter und Blüten
Salate,Gemüse
Breitblättriger Rohrkolben
junge Sprosse drei bis acht Minuten lang in Salzwasser kochen
Gemüse
Echte und HoheSchlüsselblume
innere Rosettenblätter vor der Blüte (im Frühling)
Salate, Gemüse, Suppen
Echter Dost (Oregano)
Blätter, junge Blütenstände
(Pizza-) Gewürz, Suppen
Echtes Mädesüß
Blüten
Aromatisierung von Wein, Fruchtsaft
Gänseblümchen
innere Rosettenblätter, Blütenknospen und junge Blüten
Salate, Gemüse, Suppen
Geißfuß / Giersch
junge Blätter
Salate, Gemüse, Suppen
Gemeine Braunelle (sehr häufig in Rasen und oft gemähten Wiesen)
junge Blätter
Salate, Gemüse, Suppen
Gemeine Schafgarbe
junge Blätter und Blüten
Salate, Gemüse, Suppen
Gemeiner Beinwell
junge Blätter
Salate, Gemüse, Suppen
Gewöhnlicher Beifuß
ab Juli: blütenreiche Triebe
Gewürz für fette Braten, Soßen, Suppen
Große Brennnessel
junge Blätter und Triebe
Gemüse, Suppen
Große Klette
geschälte junge Blattstiele werden wie Spargel gegessen, geschälte Wurzel
Gemüse
Gundermann
junge Triebe und Blätter
Salate, Gemüse, Suppen, Tee
Guter Heinrich
Blätter
Gemüse (Spinat-Ersatz)
Hirten-Täschelkraut
junge Blätter vor der Blüte; nicht verwenden, wenn sie durch Pilzbefall weiß gefärbt sind
Salate, Gemüse, Suppen
Huflattich
junge Blätter
Salate, Gemüse, (Rouladen)
Kleinblütiges Knopfkraut
junge Triebe
Gemüse, Suppen
Kleiner Wiesenknopf, Bibernelle
Blatter, junge Triebe
Salate, Gemüse, Suppen, Soßen, Eiergerichte
Knoblauchsrauke
nicht zu alte Blätter
Gemüse, Soßen, Suppen
Kohl-Kratzdistel
Blätter und Wurzelstock, Blütenstandsknospen
Gemüse,Wurzelstock getrocknet und gemahlen als Eindickmittel
Kriechender Günsel
Blätter, Stängel, Blüten
Salate, Gemüse, Suppen, Tee
Lungenkraut
Frühlingsblätter
Salate, Gemüse, Suppen
März-Veilchen
junge Blätter, Blüten
Salate, Gemüse, Suppen
Meerrettich
Wurzel geraspelt oder in Scheiben geschnitten
Salate, Gemüse, Suppen (sehr scharf)
Pastinak
Pfahlwurzel kann wie eine Möhre genutzt werden; Blätter und junge Sprosse
Gemüse, Suppen
Rainkohl
junge Sprosse und Blätter
Salate, Gemüse, Suppen
Sauer-Ampfer
Blätter
Salate, Gemüse, Suppen
Schmalblättriges Weidenröschen
junge Triebe (später sehr bitter!)
Gemüse
Spitz-Wegerich
junge Blätter, deren Leitbündel-fasern noch nicht zu zäh sind
Salate, Gemüse, Suppen
Thymian
Blätter, junge Blütenstände
Gewürz für Soßen, Suppen, Gemüse
Vogel-Sternmiere
Blätter; ganze Sprosse
Salate, Gemüse, Suppen
Wald-Engelwurz
junge Stängel und Blätter
Gemüse
Wasser-Minze
Blätter und Blütenstände
Tee, Salate (Würze)
Weißer Gänsefuß
Blätter
Gemüse
Wiesen-Kerbel
junge Blätter
Salate, Gemüse,Suppen
Wiesen-Schaumkraut und andere Schaumkraut-Arten
nicht zu alte Blätter, Blüten und Blütenknospen
besonders Salate (kresseartiger, scharfer Geschmack)
Wilde Malve /Weg-Malve
junge Blätter und Triebe
Salate, Gemüse, Suppen
Pflanzenfamilien
Für das Bestimmen und Wiedererkennen von Pflanzen sind die Familien, in manchen Fällen auch die Gattungen, besonders wichtig und hilfreich. Wer die in den Kästchen vorgestellten acht in der heimischen Flora häufigen und gut charakterisierten Familien wiedererkennt, wird sich mit dem Einstieg in die Artenkenntnis der Pflanzen leichter tun.
Korbblütengewächse (Asteraceae)
Vorwiegend Kräuter und Halbsträucher
Charakteristisches Merkmal sind ihre Blütenstände: Viele Einzelblüten sind zu Körbchen oder Köpfchen vereinigt, die gemeinsam oft wie eine Blüte wirken. Um die Einzelblüten zu erkennen, muss man genau hinschauen. Es gibt Blüten mit röhrenförmiger Blütenkrone und solche mit einer lang ausgezogenen Zunge. In manchen Blütenköpfchen kommen nur Röhrenblüten, in anderen nur Zungenblüten, in wieder anderen beide Blütentypen vor. Alle Blütchen haben einen unterständigen Fruchtknoten. Bei manchen besteht der Kelch aus einem Haarkranz, der auch Pappus genannt wird und später – wie beim Löwenzahn – als Flugorgan zur Verbreitung der Früchte dient.
Korbblütengewächse sind mit 21.000 Arten die artenreichste Pflanzenfamilie der Erde. Unter den mitteleuropäischen Wildpflanzen gehören zu dieser Familie so bekannte Vertreter wie Löwenzahn, Gänseblümchen, Margerite oder Flockenblumen und Disteln. Daneben zählen wichtige Salat- oder Gemüsepflanzen wie Kopfsalat, Endivie oder Schwarzwurzel und bekannte Zierpflanzen wie Aster, Zinnie, Dahlie oder Sonnenblume zu den Korbblütengewächsen. Wieder andere sind als Heilpflanzen bekannt wie Kamille, Huflattich oder Arnika.
Doldenblütengewächse (Apiaceae)
Vorwiegend Kräuter und Halbsträucher
Charakteristisch sind die fast immer als Doppeldolden ausgebildeten Blütenstände mit meist weißen, seltener gelben Blüten. Oft sind die außenstehenden Kronblätter vergrößert, sodass die ganze Doppeldolde wie eine Einzelblüte aussieht. Aus dem aus zwei Fruchtblättern zusammengesetzten Fruchtknoten bilden sich samenähnliche Früchte (z. B. Kümmelkörner oder Fenchel“samen“), bei denen Fruchtwand und Samenschale miteinander verwachsen sind). Die Blätter sind oft stark aufgeteilt (wie bei der Petersilie) und haben einen stängelumfassenden Stielgrund. Beim zerreiben kann man häufig einen aromatischen Duft wahrnehmen.
Zu der Familie gehören viele ähnlich aussehende Arten wie Wiesen-Kerbel, Kälberkropf-Arten, Wilde Möhre und Wilder Kümmel und der in Gärten wenig beliebte Giersch mit seinen ausbreitungsfreudigen unterirdischen Kriechsprossen. Gemüsepflanzen und Küchenkräuter sind Karotte (Möhre, Gelbe Rübe), Petersilie, Sellerie, Koriander, Kümmel und Kreuzkümmel. Vor allem Fenchel und Anis werden auch als Heilkräuter eingesetzt.
Lippenblütengewächse (Lamiaceae)
Vorwiegend Kräuter und Halbsträucher
Charakteristisch sind die aus fünf verwachsenen Kronblättern gebildeten zweilippigen Blüten mit nur einer Symmetrieebene. Die flache Unterlippe dient als Landeplatz für Bestäuber, die Oberlippe umschließt meist mehr oder weniger vollständig Staubblätter und Griffel. Die Sprossachsen sind meist vierkantig mit ungeteilten, gegenständigen Blättern, wobei die aufeinanderfolgenden Blattpaare um 90° gegeneinander verdreht sind („kreuzgegenständig“). Viele Lippenblütler verströmen schon bei leichter Berührung einen aromatischen Duft.
Kriechender Günsel und Gundermann blühen im Frühjahr auf Rasenflächen, wenn man sie nicht zu oft mäht, später auch die Kleine Braunelle. Taubnesseln sind häufig an Wegrändern, Wiesen-Salbei auf wenig gedüngten Wiesen, Wald-Ziest auf Wald-Lichtungen und an Waldwegen. Wegen ihrer etherischen Öle werden viele Lippenblütler als Gewürzpflanzen, Heilpflanzen oder Duftpflanzen genutzt, zum Beispiel Pfefferminze und andere Minzenarten, Majoran, Basilikum, Thymian, Bohnenkraut, Rosmarin, Salbei, und Lavendel.
Kreuzblütengewächse, Kohlgewächse (Brassicaceae)
Vorwiegend Kräuter
Der Name beschreibt die vier kreuzförmig angeordneten Kronblätter. Die Blüten sind meist in Trauben angeordnet, die oft an der Spitze sehr lange weiter wachsen. Aus den zwei Fruchtblättern entwickeln sich Schoten, wenn sie kürzer als dreimal so lang wie breit sind, spricht man von Schötchen. Die wechselständigen Blätter sind oft tief eingeschnitten, manchmal gefiedert. Häufig bildet sich eine grundständige Blattrosette aus. Typisch für Kreuzblütler ist ihr scharfer Geschmack, der auf den Gehalt an Senfölen zurückgeht und ihre Nutzung als Gewürz- und Gemüsepflanzen begründet.
Häufige Wildpflanzen sind Hirten-Täschelkraut, Hellerkraut, Schaumkraut-Arten, Hederich, Barbarakraut und Acker-Senf. Die zahlreichen verschiedenen Kohlgemüse gehen fast alle auf die Stammart Gemüse-Kohl zurück und sind der Grund dafür, dass die Kreuzblütengewächse auch Kohlgewächse genannt werden. Weitere Gewürz-, Salat- und Gemüsepflanzen sind Rettich und Radieschen, Kohlrübe, Kresse, Senf und der als Ölsaat bedeutende Raps. Abgeerntete Getreidefelder werden im Herbst oft mit Öl-Rettich oder Weißem Senf eingesät.
Das wichtigste Pflanzenmodell in der Pflanzenphysiologie und Pflanzengenetik ist das Kreuzblütengewächs Acker-Schmalwand (Arabidopsis thaliana).
Nelkengewächse (Caryophyllaceae)
Kräuter und Halbsträucher
Charakteristisch sind gabelig verzweigte Sprosse (Dichasien), bei denen in jeder Gabel eine Blüte sitzt. Die 5-zähligen Blüten sind eher flach wie bei den Mieren oder sie bilden eine lange Blütenkronenröhre wie bei den Leimkräutern oder den Nelken. Aus den Blüten bilden sich trockene Kapselfrüchte. Die Blätter sind gegenständig, meist ziemlich klein, schmal und oft ungestielt.
Mieren, Sternmieren und Hornkräuter haben kleine bis mittelgroße weiße, sternförmige Blüten. Leimkräuter haben in Platte und Nagel gegliederte freie Kronblätter, die schmalen Teile bilden in dem verwachsenen Kelch eine lange Röhre. Niederliegendes Mastkraut und Klebriges Bruchkraut sind typische Pflasterritzenpflanzen. Von der aus Südeuropa stammenden Garten-Nelke gibt es viele, meist gefüllte, weiß, rosa oder rot blühende Zuchtformen. Sie dienten oft als Bouttonière (Blume im Knopfloch des Jackenrevers).
Rosengewächse (Rosaceae)
Kräuter, Sträucher und Bäume
Blüten mit 5 Kronblättern und 5 Kelchblättern, teilweise mit Außenkelch, zahlreichen Staubblättern und ein bis vielen immer unverwachsenen Fruchtblättern. Sie entwickeln sich zu Nüsschen (bei Erdbeeren auf der fleischig verdickten Blütenachse, bei der Nelkenwurz Nüsschen mit einem Kletthaken), Sammelsteinfrüchten (Himbeere), Steinfrüchten (Kirsche) oder Apfelfrüchten. Die wechselständigen Blätter sind oft geteilt und haben am Blattgrund Nebenblätter. Bei den meisten Obstbäumen sind sie ungeteilt.
Häufige heimische Arten sind Echte Nelkenwurz, Kriechendes Fingerkraut, Wald-Erdbeeren, und als Gehölze Schlehen und Trauben-Kirschen. Wichtige Kulturpflanzen sind Obstbäume (Äpfel, Birnen, Quitten, Kirschen, Pflaumen (Zwetschgen), Mirabellen, Pfirsiche, Aprikosen) und Beeren (Erdbeeren, Himbeeren, Brombeeren). Eine häufige Gartenheckenpflanze ist die immergrüne Lorbeer-Kirsche aus Südosteuropa und dem Vorderen Orient.
Der Name geht auf die typischen Samen, die Bohnen, und auf den charakteristischen Blütenbau aus Fahne, zwei Flügeln und einem aus zwei Kronblättern verwachsenen Schiffchen zurück. Aus dem einzigen Fruchtblatt der Blüten entwickelt sich jeweils eine Hülse, weshalb die Familie auch als „Hülsenfrüchtler“ bezeichnet wird. Die Blätter sind häufig gefedert oder gefingert und haben meist am Stielgrund Nebenblätter.
Zu der Familie gehören die Klee-Arten mit dreizähligen Blättern, besonders häufig sind der Weiß-Klee und der Wiesen-Klee, und andere Gattungen, die im Gattungsnamen ebenfalls die Bezeichnung „Klee“ tragen, wie Hornklee, Steinklee, Wundklee und Schneckenklee. Weitere Gattungen sind Wicken – zum Beispiel die Zaun-Wicke und Platterbsen – zum Beispiel die Wiesen-Platterbse. Als Kulturpflanzen sind vor allem Bohnen, Erbsen, Kichererbsen und Linsen zu nennen, vorwiegend als Futterpflanzen werden Luzerne (Alfalfa) und Esparsette angepflanzt. Die weltweit wichtigste Kulturpflanze aus der Familie ist die Sojabohne. Häufige, nicht ursprünglich heimische Gehölze sind Robinie (Scheinakazie), Goldregen und die Liane Blauregen.
Lilienverwandte
Vorwiegend Kräuter
Die frühere Familie der Liliengewächse wurde vor allem aufgrund molekulargenetischer Untersuchungen in viele Familien aufgespalten. Charakteristisch für die ganze Verwandtschaftsgruppe sind sechs in zwei Wirteln angeordneten Blütenhüllblätter, die nicht in Krone und Kelch differenziert sind (Perigon), sechs Staubblätter und ein aus drei Fruchtblättern verwachsener Stempel, aus dem sich eine dreispaltige Kapsel (Tulpe) oder eine Beere (Maiglöckchen) entwickeln kann. Die Blütenhüllblätter können frei oder verwachsen sein. Die Blätter sind paralleladrig. Als unterirdische Speicherorgane kommen häufig Zwiebeln, Knollen oder Rhizome vor.
Zu den Lilienverwandten gehören Wildpflanzen wie Gelbstern, Bärlauch, Maiglöckchen und Weißwurz und viele beliebte Zierpflanzen wie Tulpen, Lilien, Hyazinthen und Kaiserkronen. Wichtige Gemüsepflanzen und Gewürze sind Küchenzwiebel, Knoblauch, Schnittlauch und Gemüselauch (Porré). Die Herbst-Zeitlose (Colchicum autumnale) enthält das gefährliche Zellgift Colchicin, das die Kernteilung hemmt.
Artenliste
(Nach Listen von Dominique Mattivi, Saskia Eberle und Jan-Niclas Frey)
Buchengallmücke (Mikiola fagi): Zitzengallen an Buchenblättern
Geißblatt-Minierfliege (Phytomyza xylostei): Gangmine im Blatt der Roten Heckenkirsche
Schaumzikade (Larve) (Philaenus spec.)
Schneeballblattkafer (Pyrrhalta viburni)
Streifenwanze (Graphosoma italicum)
Zitronenfalter (Gonepterix rhamni)
Schnecken
Rötliche Laubschnecke (Monachoides incarnatus)
Ampbibien
Gelbbauch-Unke (Bombina variegata)
Vögel/Vogelstimmen
Amsel
Buchfink
Grünspecht
Kohlmeise
Mönchsgrasmücke
Singdrossel
Zilpzalp/Weidenlaubsänger
25.Juni.2022 – In Oberteuringen und an der Rotach
Oberteuringen, ein typischer Ort Oberschwabens mit bis in die Alemannenzeit zurückgehender Siedlungsgeschichte – die erste urkundliche Erwähnung stammt von 752 –, soll zeigen, dass interessante außerschulische Lernorte auch mitten in Ortschaften zu finden sind.
Johanniskraut
Echtes Johanniskraut (Hypericum perforatum) am 24.6.2022 (Foto W.Probst)
Zum Einstieg zeige ich eine blühende Pflanze des Echten Johanniskrauts (Hypericum perforatum). Am 24. Juni war Johannis, also der Tag, der Johannes dem Täufer gewidmet ist und an dem sich Katholiken an die Taufe Jesu erinnern. Gleichzeitig ist dieser Gedenktag nahezu identisch mit der schon in vorchristlicher Zeit festlich begangenen Sommersonnenwende. Johanniskraut steht an diesem Tag in voller Blüte und wird – wie auch andere Pflanzen und Tiere, man denke an Märzenbecher, Maiglöckchen, Maikäfer oder Johanniswürmchen – nach ihrer Blütezeit oder ihren Erscheinungstermin benannt.Das Echte Johanniskraut (Hypericum perforatum) wird zudem seit altersher als Heilpflanze genutzt und seine Inhaltsstoffe, vor allem das stark färbende Antrachinon Hypericin, haben nachgewiesenermaßen beruhigende und antidepressive Wirkung. Johanniskrautöl soll auch leicht entzündungshemmende Eigenschaften haben und die Wundheilung fördern. Allerdings sind Johanniskrautpräparate nicht ohne Nebenwirkung. Die Inhaltsstoffe wirken photosensibilisierend und außerdem hemmen sie andere Medikamente zum Beispiel Herz-Kreislauf-Medikamente und die Antibabypille.
Wenn man noch geschlossene Blütenknospen zwischen Daumen und Zeigefinger presst, tritt ein durch das Hypericin rot gefärbter Tropfen aus (der symbolisch auch als Blut des Johannes gedeutet wird, der nach biblischer Überlieferung von König Herodes geköpft wurde).
Stark vergrößerter Ausschnitt aus TK 1 : 25.000, Blatt 8222, Markdorf
(1) Besondere Bäume am Sankt Martinsplatz
Die vier von uns in den Blick genommen Bäume befinden sich in einem weiter fortgeschrittenen Entwicklungsstadium. Die Purpur-Magnolie hat zwar immer noch einzelne Blüten aber auch schon relativ weit entwickelte Früchte. Catalpa steht in voller Blüte, am Judasbaum sind keine blöden oder blöden Reste mehr zu erkennen. Er hat auch keine Früchte angesetzt, vermutlich, weil es mangels eines in der Nähe stehenden Kollegen zu keiner erfolgreichen Befruchtung kam. An der Oregon-Scheizypresse fallen die zahlreichen erbsenähnlichen Zapfen auf, die schon fast ihre endgültige Größe erreicht haben, aber noch grün sind. Nähere Angaben zu den Bäumen finden sich im Bericht zur letztjährigen Exkursion.
Saponaria officinalis, das Echte Seifenkraut, liefert auch in diesem Jahr kräftigen Schaum, aber es hat nun auch schon seine zartvioletten Blüten geöffnet.
Es werden für alle Kategorien Beispielarten gefunden, aber das Sammeln und Ordnen kommt relativ früh – möglicherweise dem Klima geschuldet – zum Erliegen.
In einem schwach angestauten, flachen Bereich der Rotach suchen wir nach Wassertieren. Gefunden werden Eintagsfliegenlarven, Kleinlibellenlarven, Wasserkäfer, Flohkrebse, Plattwürmer (Strudelwürmer*) und einige Jungfische (vermutlich Flussbarsche).
*nach H. Schwab (1995): Süßwassertiere. Stuttgart: Klett der Gehörnte Vielaugen-Strudelwurm (Polycelis felina)
Mithilfe eines einfachen Auswertungssystems (Wassmann, Xylander, Naglschmid-Verlag 2014) bestimmen wir die Gewässergüte als Güteklasse II.
Das Auswertungssystem berücksichtigt den Zeigerwerte bestimmter Tiergruppen und die Formenvielfalt. Die Tiergruppen mit den empfindlichsten Tieren und die Formenvielfalt innerhalb dieser Gruppe bestimmt die „Entscheidungsklasse“, für die endgültige Bewertung ist dann außerdem die Gesamtformenvielfalt entscheidend.
Voraussetzung für die Aussagekraft ist, dass man das Gewässer relativ gründlich absucht, um die vorkommenden Arten möglichst vollständig zu erfassen. Dabei ist eine Artbestimmung nicht nötig, es sollen lediglich unterschiedlich aussehende Formen unterschieden werden. Junge (kleine) und größere (ältere) Tiere können als zwei Formen gewertet werden.
Natürlich können durch eine genauere Artbestimmung wesentlich exaktere Aussagen gemacht werden. Für die Arbeit mit Schulklassen scheint diese Methode aber sehr gut anwendbar. Unser Ergebnis stimmt mit den bei früheren Exkursionen durchgeführten aufwändigeren Untersuchungen weitgehend überein.
Der Vorteil von Gütebestimmungen mit Zeigerorganismen gegenüber chemischen Wasseranalysen liegt darin, dass man hier keine Momentaufnahme erhält, sondern eine Aussage über einen länger andauernden Zeitraum. Gerade bei stark schwankenden chemischen Werten, die zum Beispiel durch periodische Einleitungen (vor allem von Düngemittel aus der Landwirtschaft) und schwankenden Wasserständen bzw. Hochwasserereignissen zustande kommen. Dadurch ist auch die ökologische Aussagekraft besser.
(4) Nostoc, Sternenrotz oder Engelsrotz
An offenen Stellen ist der Boden mit einer schmerzlichen Kruste überzogen, die im Wasser zu olivgrünlichen, gallertigen Bändern quillt. Solche Blaugrünen Bakterien (Cyanobacteria) waren vermutlich schon vor mehr als 3 Milliarden Jahren die ersten Landbewohner. Weitere Informationen finden sich im letztjährigen Exkursionsbericht und bei Nostoc.
Das geplante Thema „Wiesen und Gräser“ muss aus Zeitmangel entfallen. Ich hoffe, wir können es in Wilhelmsdorf nachholen.
Der Schutz und die Wiederherstellung von Mooren gilt schon seit langem als wichtige Naturschutzaufgabe. Dabei ging es zunächst in erster Linie um die schützenswerten Lebensgemeinschaften mit ganz besonderen, in der übrigen Landschaft seltenen oder fehlenden Arten. Erst durch die hohe Aktualität der Klimakrise rückte die Bedeutung der Moore als Kohlenstoffspeicher in den Vordergrund. Aber auch ihre Bedeutung für den Wasserhaushalt und den Stickstoffkreislauf befördert aktuelle Moorschutzmaßnahmen.
Feuchtbiotope
Unter Feuchtbiotopen versteht man Lebensraumtypen, die über einen längeren Zeitraum des Jahres bis zur Landoberfläche mit Wasser gesättigt sind. Weiter gefasst werden auch Seen und Fließgewässer und von Salzwasser bestimmte Lebensräume wie das Wattenmeer mit einbezogen. Obwohl solche Feuchtgebiete nur etwa 6 % der Erdoberfläche einnehmen, erbringen sie rund ein Viertel der Nettoprimärproduktion. Sie haben eine besondere Bedeutung als Grundwasserfilter, für Überschwemmungsschutz, in vielen Fällen als Kohlenstoffsenke und als Rast- und Überwindungsplätze für Wasser- und Watvögel.
Man unterscheidet zum Beispiel Moore, Brüche, Auwälder, Riede und Sümpfe. Für die Einteilung ist wichtig, ob Torfbildung stattfindet oder nicht und wie die Wasserversorgung des Gebietes erfolgt. Auch das Vorhandensein oder Fehlen von Bäumen und anderen Gehölzen spielt für die Unterscheidung eine wichtige Rolle.
Abb. 1 Überblick über die verschiedenen Feuchtbiotope in Mitteleuropa
Moore als Kohlenstoffspeicher
Für die Kohlenstoffspeicherung von besonderer Bedeutung sind Moore. Sie entstehen auf wasserdurchtränkten Böden, in denen wegen des Sauerstoffmangels die anfallenden Pflanzenreste nur sehr langsam zersetzt werden. Da die Produktion von organischer Substanz rascher erfolgt als ihr Abbau, kommt es zur Ablagerung von Torf. Dabei ist „Moor“ ein geografischer bzw. botanischer, „Torf“ ein mineralogisch-petrografischer Begriff. Bodenkundlich ist Torf definiert durch seinen hohen Glühverlust (bei 550 °C):Torf: 100-75 %, anmooriger Boden: 74-15 %, Mineralboden: unter 15 %.
Wenn Torfschichten eine Mächtigkeit von über 30 cm haben werden diese Gebiete als Moore bezeichnet, unabhängig davon, ob dort noch eine neue Torfbildung stattfindet oder nicht. Bei einer geringeren Torfschicht oder einem geringeren Torfanteil im Boden spricht man von „Anmoor“. Der Überbegriff für beide ist „organische Böden“. Im Gegensatz dazu haben mineralische Böden einen geringeren organischen (Humus-)Anteil und einen höheren Anteil aus verwittertem Gestein.
Beim Abbau der organischen Substanz unterscheidet man:
Verwesung durch aerobe Mikroorganismen: Völliger Abbau zu Kohlenstoffdioxid und Wasser sowie anorganischen Mineralstoffen (Nitrate, Phosphate….).
Vermoderung: Unvollkommene Verwesung bei unzureichendem Sauerstoffzutritt.
Fäulnis: Vollzieht sich unter Sauerstoffabschluss; es bilden sich durch anaerobe Bakterien vor allem Methan und Schwefelwasserstoff, aber auch Ammoniak und Lachgas; Bildung von Faulschlamm, Mudde (Seesediment mit relativ hohem organischem Anteil).
Vertorfung beginnt bei behindertem Sauerstoffzutritt mit Vermoderung, später folgt unter Luftabschluss eine sehr langsame Fäulnis. Schnell zersetzen sich die Zellinhalte aus Proteinen, Zuckern und Stärke. Langsamer werden die Stoffe der Zellwände abgebaut, zuerst Pektine und Hemizellulosen, dann die Zellulose zuletzt der Holzstoff Lignin. Sehr schwer zersetzen sich außerdem Fette, Harze,Wachse, Kutin und Sporopollenin. Pollenkörner und Sporen bleiben in Torf deshalb sehr gut erhalten. Durch ihre Funde in gut datierbaren Torfschichten kann man deshalb auf die Vegetation früherer Zeiten schließen (Pollendiagramme).
Abb. 2 Torfbildung
Für die Eigenschaften des Torfes (Struktur, Anteil an Mineralstoffen, Huminstoffen, pH-Wert, Wassergehalt) ist die Pflanzengemeinschaft wichtig, aus deren Ablagerungen er entstanden ist. Immer handelt es sich dabei um Pflanzengemeinschaften feuchter Standorte.
Abb. 3 Torfbildung von unterschiedlichen Pflazengemeinschaften (verändert nach Overbeck 1975)
Die Anhäufung von organischem Material in aktiven Mooren ist standortabhängig. Aus Messungen ergibt sich ein Torfwachstum von 1± 0,8mm im Jahr. Die großen Unterschiede kommen durch die unterschiedliche torfbildende Vegetation und die klimatischen Bedingungen zustande.
In jedem Fall wird der Atmosphäre solange Kohlenstoff entzogen, solange mehr Torf gebildet als abgebaut wird. Moore gelten daher als Kohlenstoffsenken. Für die langfristige Kohlenstoffakkumulation unterschiedlicher Torfarten hat man Werte zwischen 0,15 und 1,3 t C ha-1 a-1 ermittelt (Tepel 2007/08). Das unterscheidet Moore von Wäldern, deren Senkenwirkung mit dem Erreichen des Klimaxstadiums beendet ist, da sich dann Einlagerung und Abgabe die Waage halten. Aber auch trockengelegte, kultivierte oder anderweitig genutzte Moore können von Kohlenstoffsenken zu Kohlenstoffquellen werden, da ihr Kohlenstoffspeicher durch aerobe oder anaerobe Zersetzungsvorgänge abgebaut wird. Bei aerobem Abbau wird Kohlenstoffdioxid, bei anaerobem Methan freigesetzt. In ausgetrockneten Mooren wird dies in den oberen Schichten jedoch schnell zu CO2 oxidiert (Abb. 4). Durch Vernässung kann die Torfbildung wieder in Gang gebracht und damit die Wirkung als Kohlenstoffsenke wiederhergestellt werden.
Abb.4 Moore als Kohlenstoffsenken und -quellen
Etwa 3 % der Landfläche der Erde sind von Mooren oder Anmooren bedeckt. Das entspricht einer Fläche von 4 Millionen km². Die größten Moorflächen finden sich in Kanada, Alaska, Nordeuropa und Sibirien, aber auch in tropischen Waldgebieten von Südostasien, im Amazonasbecken und im Kongo-Regenwald wurden große Torfflächen nachgewiesen (Page/Rieley/Wüst 2006, Dargie et al. 2017). In Mitteleuropa sind ursprünglich etwa 5 % der Landfläche von Mooren bedeckt. Sie sind alle nach der Eiszeit beginnend vor etwa 15.000 Jahren entstanden und zwar in den von Gletschern überformten Gebieten Norddeutschlands und am Alpenrand. Einige Moore gibt es auch in den Mittelgebirgsräumen, beispielsweise im Hohen Venn und im Schwarzwald.
Tab.1 Aufteilung der Landfläche auf der Erde (2019) (nach Jäger 2020)
Fläche in106 km2
Anteil an der Landfläche in %
gesamte Landfläche
149
landwirtschaftlich genutzte Fläche
51
34
Wälder
39
26
Gletscher, Wüsten u.Ä.
43
29
Busch
12
8
Siedlungen
1,5
1
Seen, Flüsse
1,5
1
in den genannten Flächen enthalten:
Moore und Anmoore (organische Böden).
ca.4
3
Tab.1 Aufteilung der Landfläche auf der Erde (2019) (nach Jäger 2020)
Global ist die Menge an organisch gebundenen Kohlenstoff in den Böden ungefähr dreimal so groß wie die Kohlenstoffmenge in allen Lebewesen zusammen und doppelt so groß wie der Kohlenstoffgehalt der Atmosphäre.
System
Kohlenstoffvorrat (in Gt)
Böden insgesamt
1500
Moorböden
ca.500
Landpflanzen
560
Atmosphäre
750
Ozeane
38.000
Marines Plankton
3
Tab. 2 Kohlenstoffvorräte in Gigatonnen für unterschiedliche Systemkompartimente des Kohlenstoffkreislaufs (nach Trepel 2007/08). Dank des mittlerweile (2022) auf 416 Vol ppm angestiegenen CO2-Gehalts der Atmosphär beträgt der Kohlenstoffvorrat derzeit ca. 850 Gt.
Nach einer Datenauswertung von Yu et al. von 2010 zeigt sich, dass die Kohlenstoffspeicherung nach der letzten Kaltzeit in den Mooren der Nordhemisphäre am höchsten war, wobei höchste Akkumulation im frühen Holozän lag. Deutlich weniger Kohlenstoff wurde in tropischen Moorgebieten vor allem vor 4000-8000 Jahren akkumuliert, während die Moore der Südhemisphäre – vor allem in Patagonien gelegen – vor allem während einer Wärmeperiode vor 15-20.000 Jahren Torfschichten aufgebaut haben
Region
Fläche (km2)
C-Speicher(Gt)
durchschnittliche C-Speicherung (gCm-2a-1) seit der letzten Vereisung
Nordhemisphäre
4 000 000
547 (473-621)
18,6
Tropen
368 000
50(44-55).
12,8
Südhemisphäre
45 000
15 (13-18)
22,0
Tab. 3 Überblick über die Moorflächen der Erde und ihre Kohlenstoffspeicherung (nach Yu et al. 2010)
Fläche in ha
Gespeicherte Kohlenstoff in G t
Organische Böden in der EU
31 000 000
17
Organische Böden in Deutschland
1 823 922
mindestens 1,3
Tab. 4 Organische Böden in Europa und ihre Kohlenstoffspeicherung (nach Jäger 2020)
Für die Klimaerwärmung spielt vor allem die Vernichtung von Kohlenstoffvorräten in den Moorböden weltweit eine wichtige Rolle. Torfbrände in Südostasien haben zum Beispiel in den letzten Jahrzehnten den stärksten Anstieg der CO2-Emissionen in der Atmosphäre bewirkt (Page et al 2002, Rieley et al. 2006). In Deutschland spielt vor allem die landwirtschaftliche Nutzung von Moorböden eine entscheidende Rolle für die Freisetzung von Kohlenstoffdioxid.
Bereiche
in Mt CO2– Äquivalente pro Jahr
aus allen Bereichen in Deutschland
ca. 900
aus Landwirtschaft (ohne die Herstellung synthetischer Düngemittel)
103,5
aus organischen Böden, die als Acker und Grünland genutzt werden
38
Tab. 5 Treibhausgasemissionen in Deutschland (nach Jäger 2020)
Moortypen und ihre Entstehung
Je nach Umweltbedingungen entstehen unterschiedliche Moortypen. Sie unterscheiden sich vor allem darin, woher das Wasser kommt, welche Salze im Wasser gelöst sind und welche Pflanzenarten deshalb dort gedeihen können. So werden die regenwasserabhängigen Hochmoore oder Regenmoore den Niedermooren gegenübergestellt, die ihren Wasservorrat aus dem Grundwasser oder aus Oberflächengewässern erhalten. Regenwasser ist sehr mineralstoffarm. Der Mineralstoffgehalt der Gewässer, die Niedermoore speisen, kann sehr unterschiedlich sein. Nach der Herkunft des Wassers kann man sehr verschiedene Niedermoortypen unterscheiden.
Niedermoore (Wasserversorgung durch Oberflächenabfluss und Grundwasser)
Verlandungsmoore
Versumpfungsmoore
Überrieselungsmoore, Durchströmungsmoore
Quellmoore
Flussüberflutungsmoore
Niedermoore können je nach Nährmineralien und Kalkgehalt zahlreiche seltene Pflanzenarten beherbergen, zum Beispiel Seggen-Arten und Orchideen.
Hochmoore (Wasserversorgung nur durch die Niederschläge)
allmählich aus mineralstoffarmem Niedermoor (über Verlandung oder Versumpfung)
direkt (Wurzelechtes Hochmoor) auf feuchtem, nährmineralarmen Böden
Abb. 5 Moortypen
Hochmoore
Aufbau und Hochmoortypen
Abb. 6 Aufbau eines mitteleuropäischen Hochmoors
Das Aussehen und der Aufbau der Regenmoore verändert sich von dem sehr atlantischen Klima des äußersten Westeuropas zum kontinentalen Klima Osteuropas. Die Deckenmoore Schottlands und Irlands haben sich aus ursprünglich bewaldeten Gebieten durch menschlichen Einfluss, insbesondere durch Beweidung, an waldfreien Standorten entwickeln können.
Abb. 7 Aussehen der Regenmoore in unterschiedlichen Klimabereichen Europas
Nach Norden schließt an die Zone der echten Hochmoore die Zone der Aapamoore an. Sie sind im kalt gemäßigten Klima zirkumpolar verbreitet und bestehen aus hangparallel verlaufenden Wällen und Senken. Die Wälle haben Hochmoorcharakter (ombrotroph), die Senken Niedermoorcharakter (minerotroph). Noch weiter nach Norden, nördlich der Baumgrenze, folgt die Zone der Palsenmoore, deren hügelartige Strukturen an mehrjähriges Bodeneis gebunden sind. Noch weiter nach Norden folgen auf durchgehend gefrorenen Permafrostböden Polygonmoore, deren polygonartige Strukturen durch Frosttrockniss entstanden sind, als nach einer längeren Feuchtperiode im Atlantikum (7270-3710 v. Chr.) das Klima kälter wurde. Dieser Moortyp ist typisch für Nordostsibirien und er ist besonders vom Klimawandel bedroht (POLYGON, Uni Greifswald 2011-2014).
Abb.8 Nördliche Moore
Torfmoose und Hochmoorwachstum
Voraussetzung für die Hochmoorbildung ist die Ansiedlung von Torfmoosen (Gattung Sphagnum).Torfmoose können aufgrund ihres anatomischen Baus das 20 bis 30 fache ihres Trockengewichtes an Wasser aufnehmen und speichern. Außerdem gestattet ihnen ein besonderer Ionenaustauschmechanismus selbst aus extrem nährmineralarmen Wasser die wenigen enthaltenen Kationen im Austausch gegen H+– Ionen herauszufangen. Dies bewirkt eine sehr starke Ansäuerung des Wassers (bis zu pH 3 (Dierßen u. Dierßen 2008) und damit eine weitgehende Ausschaltung von Konkurrenten. Als Ionenaustauscher wirken dabei vor allem bestimmte Substanzen in der Zellwand. Ob die so herausgefangenen Ionen tatsächlich der Mineralstoffzufuhr der Sphagnum-Pflanze dienen, ist allerdings fraglich.. Möglicherweise ist entscheidend, dass auf diese Weise für die Sphagnumzellen giftige Calcium- und Aluminiumionen aus dem aufsteigenden Wasser entfernt werden.
Abb. 9 Morphologie der Torfmoose (Sphagum magellanicum)Abb. 10 Räumliche Darstellung eines Sphagnum-Blättchens mit toten Hyalocyten ( Wasserspeicherzellen) und lebenden Chlorocyten
Die Torfmoospolster und – decken wachsen immer höher über den Grundwasserspiegel hinaus und in dem abgestorbenen Moostorf hält sich das Regenwasser wie in einem Schwamm. So können bis zu 5 m über das Relief emporgewölbte Torfschilde entstehen, aus denen am Rand ständig saures, nährsalzarmes Wasser abfließt und sich über das Randgehänge in dem sogenannten Randsumpf („Lagg“) ansammelt. Dieser Randsumpf ist dadurch etwas mineralstoffreicher als die Moorhochfläche.
Dabei wächst die Torfmoosdecke nicht gleichmäßig in die Höhe. Man unterscheidet zwischen höheren Bulten und tieferen Schlenken. In den Schlenken ist der Zuwachs am stärksten, dadurch werden aus Schlenken mit der Zeit Bulte und umgekehrt.
Abb. 11 Bult-Schenken-Komplex (Abbildung aus Probst, W. 1978)
In vielen Veröffentlichungen wird angegeben, dass das Torfwachstum in Mitteleuropa etwa 10 cm pro 100 Jahre beträgt. Die größten Torfmächtigkeiten, die man erbohrt hat, liegen um 10 m. Dies würde einer Entstehung unmittelbar nach dem Ende der Eiszeit entsprechen. Allerdings sind die Wachstumsraten – wie schon oben ausgeführt – stark von den jeweiligen Umweltbedingungen abhängig. Außerdem kann man davon ausgehen, dass sich das Hochmoorwachstum mit zunehmender Höhe verlangsamt, da sich der schwerkraftbedingte Wasserabfluss verstärkt und außerdem Zersetzungsvorgänge in den tieferen Schichten und zunehmender Druck der darüberliegenden Schichten zu einem Zusammensacken führen.
In dem obersten halben Meter eines Hochmoores lässt sich ein Torfbildungshorizont (Akrotelm, von lat. telma = Moor) von einem Torfablagerungshorizont (Katotelm) unterscheiden. In einer obersten etwa 2-5 cm dicken Schicht des Akrotelms sind die Torfmoose photosynthetisch aktiv (euphotische Zone). An der Untergrenze dieser Schicht beträgt die Lichtintensität noch etwa 1 % des Oberflächenwertes. In der anschließenden aphotischen Zone, einer 10-50 cm dicken Schicht, sind die Torfmoose weitgehend abgestorben. Sie ist noch von lebenden Wurzeln der Gefäßpflanzen durchzogen. Abgestorbene Pflanzenteile werden von Bakterien und vor allem von Pilzen aerob abgebaut. Der Stickstoffgehalt ist hier noch niedriger als in der Oberflächenschicht (C/N bis 75 gegenüber C/N 50 in der Wachstumszone der Torfmoose, Dierßen und Dierßen 2008).
Unterhalb der aphotischen, noch sauerstoffhaltigen Zone folgt das Katotelm, beginnend mit einer Verdichtungszone von 2-15 cm Mächtigkeit. Die Pflanzenreste sind hier schon stärker zersetzt und werden durch das aufliegende Gewicht verdichtet. Darunter folgt ein mehr oder weniger ausgedehntes Torflager. Wegen der starken Verdichtung ist es nur wenig wasserdurchlässig. Der im Wasser enthaltene Sauerstoff ist deshalb schnell verbraucht und die weiteren Zersetzungsvorgänge werden nun von Anaerobiern übernommen, wobei vor allem Methan gebildet wird .
Abb. 12 Hochmoorschichtung
Abb. 13 Sumpf-Torfmoos (Sphagnum palustre). Der Übergang von der euphotischen in die aphotische ist gut an der Farbänderung zu erkennen.
Aus der weiteren Schichtenfolge lässt sich die Entstehungsgeschichte des Moores ableiten. In der Abbildung ist die Schichtenfolge in einem Verlandungs-Hochmoor dargestellt.
Abb. 14 Schichtenfolge in einem Verlandungs-Hochmoor
Das Torfmoos-Mikrobiom und mögliche symbiotische Beziehungen
Die Erforschung des Mikrobioms der Sphagnumpflanzen ist noch in ihren Anfängen und erst durch neueste Möglichkeiten der Genomsequenzierung (next generation sequencing) wurden Fortschritte erzielt. Zunächst ging es um den Nachweis der verschiedenen beteiligten Mikrobionten. In den Sphagnumpflanzen befinden sie sich vor allem in den wasserspeichernden Hyalocyten, in den lebenden Chlorocyten konnten nur wenige Bakterien nachgewiesen werden. Man kann die Hyalocyten geradezu als kleine Kulturgefäße für Mikroben ansehen, von denen die Moose profitieren. Wie Untersuchungen an lebenden Sphagnumköpfchen zeigten, enthalten sie vor allem Proteobakterien und Acidobakterien. Cyanobakterien und Archäen spielen kaum eine Rolle (Kostka et al. 2016).
Untersuchungen zur Funktion des Mikrobioms ergaben eine besondere Bedeutung methanotropher Proteobakterien, die gleichzeitig azidotroph sind, also N2 assimilieren. Dies könnte erklären, warum die Stickstoffspeicherung in Sphagnummooren in Gebieten mit sehr geringen Konzentrationen von Stickstoffverbindungen in der Luft deutlich höher ist als der daraus zu erwartende Stickstoffgehalt. Das „Futter“ für die methanotrophen Bakterien liefert das in tieferen Moorschichten von methanogenen Bakterien und Archäen produzierte Methan. Der Sauerstoff wird auch von den Photosynthese betreibenden Sphagnumköpfchen bereitgestellt. Möglicherweise könnten die Bakterien auch von den Torfmoos-Chlorocyten abgegebenen Kohlenhydraten profitieren. Durch Isotopmarkierung konnte nachgewiesen werden, dass sich der Luftstickstoff tatsächlich in Proteinverbindungen der Sphagnen wieder finden lässt (Vile et al. 2014). Dorthin könnte er durch direkte Abgabe von Stickstoffverbindungen (zum Beispiel Ammonium) durch die methanotrophen Bakterien oder über die Freisetzung von Stickstoffverbindungen aus abgestorbenen Bakterien gelangt sein. Auch Konsumenten der Bakterien könnten die Sphagnen über ihre Ausscheidungen düngen. Die Hinweise verdichten sich, dass es sich bei diesen Stoffwechselbeziehungen um eine echte Symbiose handelt, vergleichbar mit Knöllchenbakterien und Leguminosen.
Abb.15 Mögliche Stoffumsätze in der obersten Torfmoosschicht. Zwischen Sphagnen und methanotrophen Proteobakterien besteht eine symbiotische Beziehung.
Abb. 16 Beziehungen zwischen Sphagnum und methanotrophen Proteobakterien
Es wäre denkbar, dass ein erhöhter Eintrag von Stickstoffverbindungen aus der Luft zu einer Verringerung der N2 Assimilation führen würde. Dies könnte wiederum die Methanabgabe der Moore beeinflussen (erhöhen) (Vile et al. 2014).
Pflanzen und Tiere
Auf wachsenden Hochmoorflächen kommen nur wenige Gefäßpflanzenarten vor. Neben dem Scheidigen Wollgras (Eriophorum vaginatum, vgl. Titelbild) sind dies die Heidekrautgewächse Moosbeere (Vaccinium oxycoccus) und Rosmarinheide (Andromeda polyfolia) sowie der insektenfressende Rundblättrige Sonnentau (Drosera rotundifolia). An trockeneren Bereichen können sich als weitere Heidekrautgewächse Gewöhnliche Glockenheide (Erica vulgaris) und Besenheide (Calluna vulgaris) ansiedeln, im Randbereich auch Heidelbeeren (Vaccinium myrtyllus), Preiselbeeren (Vaccinium vitis-idaea) und Rauschbeeren (Vacciinium uliginosum), in von atlantischem Klima geprägten Bereichen Norddeutschlands auch der Gagelstrauch (Myrica gale) und die Krähenbeere (Empetrum nigrum), in Bereichen mit etwas kontinentalerem Klima Nordostdeutschlands der in Deutschland sehr selten gewordene Sumpf-Porst (Rhododendron tomentosum, Syn.:Ledum palustre). Weitere Hochmoorpflanzen sind In feuchteren Bereichen das Weiße Schnabelried (Rhynchospoa alba), Schmalblättriges Wollgras (Eriophorum angustifolium) und weitere Zypergrasgewächse.
Abb. 17 Beispiele für Gefäßpflanzen des Hochmoors
Auch die Fauna der Hochmoore besteht vorwiegend aus Spezialisten. Für Fische ist das Wasser zu sauer, wegen des Calciummangels fehlen Schnecken, Muscheln und Krebse. Typische Hochmoor-Insekten sind zum Beispiel die Hochmoor-Mosaikjungfer (Aeschna subarctica) und der Hochmoor-Perlmutterfalter (Boloria aquilonaris), dessen Raupe sich von Moosbeeren ernährt. Unter den Wirbeltieren sind vor allem der Moorfrosch und die Kreuzotter – oft in ihrer schwarzen Variante – zu nennen Regelmäßig in Hochmooren anzutreffende Vögel sind zum Beispiel Großer Brachvogel, Goldregenpfeifer, Kranich, Birkhuhn, Sumpfohreule, Krick – und Knäkente.
Tropische Moore
Torfbildung findet vor allem in kühleren Klimaregionen statt, wo der Abbau organischer Substanz insgesamt langsamer verläuft. Aber es gibt auch Torfgebiete unter tropischen Sumpfwäldern, zum Beispiel im Amazonasgebiet, im Kongobecken und in Indonesien. Voraussetzung sind hohe Niederschläge – deutlich über 2000mm im Jahr – welche die Evaporation übersteigen.
Die großen Torflagerstätten in der zentralen Senke des Kongobeckens, der sogenannten Cuvette Centrale, wurden erst vor wenigen Jahren entdeckt und vermessen. Die Torfschichten sind zwischen 2,4 und 5,9 m dick (Dargie et al. 2022). Die Wissenschaftler stellten fest, dass die Torflager immer unter bestimmten Waldgesellschaften auftreten, deren Ausdehnung sie mithilfe von Satellitenbildern auf 145.000 km² berechnen konnten. Das sind knapp 10 % des gesamten Kongobeckens. Nach Berechnungen der Forscher könnten in diesem Torflager 30,6 Milliarden t Kohlenstoff gespeichert sein.
Die Fläche der Moorgebiete in Südostasien wird auf 230.000 km² geschätzt (Page, Riley, Wüst 2006). Sie sind stark bedroht durch Brandrodung und Umwandlung in Agrarflächen. In unberührten Zustand haben diese Moore einen niedrigen pH-Wert (3-4) und niedrige Nährmineraliengehalte. Der Gehalt an organischem Kohlenstoff übertrifft 50 %, während der Stickstoffgehalt bei 2 % liegt. Im Gegensatz zu nördlichen Hochmooren ist der Ligningehalt des Torfes hoch und der Zellulosegehalt relativ niedrig. Dies hängt damit zusammen, dass die Vegetation dieser tropischen Moore vor allem aus Gehölzen besteht. Ihre Kohlenstoffspeicherung wird auf 50-70 Gigatonnen geschätzt, der jährliche Zuwachs ist unter günstigen Bedingungen drei bis viermal so hoch wie bei nördlichen Regenwassermooren.
Mensch und Moor
Brennstoff
. In Irland, Finnland und Schweden gibt es bis heute Stromkraftwerke, die mit Torf betrieben werden. Früher wurden die in Ziegelform gebrachten Torfbriketts an der Luft getrocknet, bevor sie als Brennmaterial genutzt werden konnten. In manchen Mooren wurden die Flächen kleinparzellig aufgeteilt, und die einzelnen Parzellen wurden von unterschiedlichen Landwirten zur Brennstoffgewinnung genutzt. Aus den kleinen Torfstichen solcher Moore ist – bei mäßiger Entwässerung – eine Regeneration möglich.
Abb. 18 Besitzverhältnisse im Jardelunder Moor bei Flensburg (Katasterplankarte 1:5000, Stand 1978)
Braunkohle und Steinkohle sind fossile Torfe.
Gartenbau
Heute dient der Torfabbau vor allem der Gewinnung von Pflanzensubstrat in der Gärtnerei, für Presstöpfe zur Sämlingsanzucht und für Wurzelballen der meisten im Handel angebotenen Pflanzen, sowie für die meisten käuflichen Blumenerden. Im Gegensatz zum Brennmaterial ist zu diesem Zweck Weißtorf besonders gut geeignet. Es handelt sich um ein sehr einheitliches Substrat mit ausgezeichneter Wasseraufnahmefähigkeit und der Fähigkeit zur Mineralstoffspeicherung. Sein niedriger pH-Wert kann durch Kalkung bis über den Neutralpunkt hinaus verändert werden. So können mit diesem Grundsubstrat sehr unterschiedliche Pflanzsubstrate hergestellt werden.
2018 wurden in Deutschland etwa 3,7 Millionen m³ Torf abgebaut – von 2002-2009 waren es nach Auskunft der Bundesregierung noch durchschnittlich 8,2 Millionen m³ pro Jahr – und rund 4,1 Millionen m³ importiert, vor allem aus dem Baltikum. Allerdings wurden in Deutschland seit den 1980er Jahren keine intakten Moore mehr für den Abbau freigegeben, sondern nur noch Gebiete, die vorher landwirtschaftlich genutzt wurden. Die zu entnehmenden Torfmengen werden genau vorgegeben und es besteht eine Renaturierungspflicht für die Abbauer (Bundesinformationszentrum Landwirtschaft 2020). Alte Abbauverträge sind davon allerdings nicht berührt (s.u. Reichermoos) .
Ein völliger Verzicht von Torf im Erwerbsgartenbau wäre prinzipiell möglich aber sehr aufwendig, denn alle Ersatzsubstrate haben keine so guten und einheitlichen Eigenschaften wie Hochmoortorf. Infrage kommen Grünkompost, Rindenhumus Holzfasern. Kokosfasern, Blähton oder Perlit (Amberger-Ochsenbauer, Meinken 2020).
Medizin
Für Medizin und Körperpflege spielen Moorbäder und Moor-(Fango) packungen (von lat. fango = Schlamm, Schlick) eine wichtige Rolle. Der dickflüssige Brei aus Schwarztorf wird mit Temperaturen von 38-40° verwendet. Neben der Wärme sollen vor allem die im Torf enthaltenen Huminsäuren nicht nur die Haut weich machen und die Durchblutung fördern, sondern auch eine günstige Wirkung auf das endokrine System ausüben.
Abtorfung im Reichermoos bei Vogt, Kreis Ravensburg. Für die Heilbäder in Bad Wurzach, Bad Waldsee, und Bad Buchau soll dieser Torf nach der Regionalplanung von 2021bis zum Jahr 2070 sich abgebaut werden. 1970 verpachtete das Land Baden-Württemberg den Torfabbau im Reicher Moos. Die Pächter fräßen der Torf mit riesigen Maschinen ab. Dagegen wendet sich eine Bürgerinitiative. (Foto W. Probst, 5.7.1983)
Filtermaterial
In der Aquaristik und in der Teichwirtschaft wird Torf als Filtermaterial zur Herabsetzung des pH-Wertes und der Carbonathärte verwendet. Außerdem sollen die Fulvosäuren im Schwarztorf die Schleimhäute der Fische vor bakteriellen Infektionen schützen. Durch Torffilterung kann man das Aquarienwasser den Verhältnissen in tropischen Schwarzwasserflüssen annähern, aus denen viele Zierfische stammen. Als natürlicher Ionenaustauscher kommt Torf auch in der chemischen Industrie zum Einsatz. Aus Torf lässt sich auch Aktivkohle zur Filterung herstellen, die vor allem in Chemielabors zum Einsatz kommt.
Weitere Nutzungen
Torffasern eignet sich zur Herstellung von Isolationsmaterial, sie lassen sich zu leichten und warmen Textilien und Unterlagen verarbeiten. Bis heute dienen Torffasern als natürlicher Füllstoff für Matratzen, Bettdecken und Kissen.
Vor allem im Pferdeställen wurde Torf als Einstreu genutzt.
Moorkultivierung
Die großen Moorflächen vor allem in Norddeutschland aber auch im süddeutschen Alpenvorland waren lange Zeit landwirtschaftlich nicht zu nutzen. Um die Ernährung der wachsenden Bevölkerung sicherzustellen, wurden deshalb immer wieder Versuche unternommen solche Moorflächen für die landwirtschaftliche Produktion nutzbar zu machen.
Die sogenannte Fehnkultur (von niederländisch Veen = Moor) wurde in den Niederlanden entwickelt aber schon im 17. Jahrhundert auch in Nordwestdeutschland angewandt. Dabei wurden zunächst tiefe Entwässerungskanäle angelegt, durch die der gestochene Torf mit Schiffen abtransportiert werden konnte. Auf dem Rückweg wurde von den Schiffen dann Schlick mitgebracht und vor allem mit dem Weißtorf vermischt. Beidseitig der Kanäle entstanden nach und nach typische Fehnsiedlungen.
Vor allem Im Laufe des 18. und 19. Jahrhunderts wurden in Deutschland verschiedene weitere Arten der Moorkultivierug entwickelt. Dabei spielten Entwässerung, Abtorfen, Brennen, Tiefpflügen zur Vermischung mit dem mineralischen Untergrund und Kalkdüngung eine wichtige Rolle. Oft wurde die schwierige Bearbeitung der Torfböden durch neue Siedler geleistet, die aus ihrer Heimat durch Not oder Verfolgung vertrieben worden waren.
Alle Kultivierungsmaßnahmen führten dazu, dass die Torfneubildung und -ablagerung gestoppt wurde und dadurch aus der Kohlenstoffsenke durch anaeroben Abbau der Torfschichten eine Kohlenstoffquelle wurde.
Paludikultur
Eine neue Form der Moornutzung ist die „Paludikultur„. Kulturpflanzen sind hier die Torfmoose, die großflächig unter Hochmoorbedingungen kultiviert werden. Die Torfmoosernte soll den Torfabbau ersetzen. Dadurch wird die Kohlenstofffreisetzung der üblichen Moorkultivierung verhindert und eine ökonomisch tragbare Alternative aufgezeigt. Nasskulturen können außer auf Hochmoorstandorten auch auf Nieder- und Zwischenmooren und anderen kohlenstoffspeichernden Feuchtgebieten entwickelt werden. Die produzierte Biomasse aus Schilf, Binsen, Sauergräsern und anderen Feuchtpflanzen könnte als Material für unterschiedliche Baustoffe verwendet werden (Wichtmann, Schröder, Joosten, 2016).
Möglichkeiten des Moorschutzes
Nach Dierßen und Dierßen (2008) gibt es im Prinzip drei Möglichkeiten des Schutzes:
Bewahren eines derzeitigen Zustandes bzw. zulassen einer natürlichen Sukzession ohne Eingriffe
Pflegen eines aktuellen wünschenswerten Zustandes
Entwickeln eines Zustandes, der den jetzigen Zustand verbessert, durch geplante Pflege und Steuerungseingriffe (Restitution)
Die erste Vorgehensweise bietet sich an, wenn der derzeitigen Zustand sehr gut ist und sich durch Eingriffe kaum verbessern lässt oder wenn man erwarten kann, dass eine natürliche Sukzession zu einem wünschenswerten Zustand führt. Ein intaktes Hochmoor mit funktionierendem Bult-Schlenken-Komplex sollte vor Eingriffen abgeschirmt werden. Aber auch ein teilweise abgetorftes Hochmoor, bei dem sich in Torfstichen gute Sukzessionen mit Torfmoosen entwickeln, kann man am besten sich selber überlassen.
In vielen Fällen kann man erkennen, dass ein derzeitiger guter Zustand dabei ist, sich zu verschlechtern. So können noch vorhandene Bult-Schlenken-Komplexe bei zunehmender Austrocknung immer stärker von Besenheide besiedelt werden und ihr Wachstum einstellen. In diesem Fall könnten Maßnahmen gegen die Entwässerung und Austrocknung den besseren Zustand erhalten. Auch das starke Aufkommen von Baumwuchs, vor allem von Birken, ebenfalls im Zusammenhang mit Austrocknung aber auch mit Nährmineraleintrag, kann durch Entfernen des Birkenaufwuchses gebremst werden. In jedem Fall ist bei allen Maßnahmen eine gründliche Analyse der Wirkungszusammenhänge Voraussetzung für einen Erfolg.
Besonders schwierig ist die Restitution, im Hinblick auf Hochmoore also die Entwicklung relativ nährmineralreicher und von menschlichen Aktivitäten stark beeinflusster Flächen zurück zu nährmineralarmen, vom Regenwasser abhängigen Torfmoosflächen. Dies liegt vor allem daran, dass sich in der von Landwirtschaft, Siedlungen und Verkehr geprägten mitteleuropäischen Kulturlandschaft Düngemitteleintrag und Entwässerung kaum vermeiden lassen.
Abb. 19 Wiedervernässte Fläche im Wurzacher Ried
Moore im Biologieunterricht
Mögliche Unterrichtsthemen
Vom Gletschersee zum Hochmoor – ein Beispiel für nacheiszeitliche Landschaftsentwicklung
Für einige mitteleuropäische Moore ist die Entwicklung vom Eisstausee am Ende der letzten Kaltzeit bis zum Hochmoor gut dokumentiert. Diese zeitliche Entwicklung lässt sich bei einer Reise in den Untergrund nachvollziehen.
Abb. 20 Mit den verschiedenen Sedimentschichten eines Moores kann man in die Vergangenheit reisen
Speicher, Senken, Quellen? – Wie Moore sich auf die Treibhausgase der Atmosphäre auswirken
Der aus wenig zersetzen pflanzlichen Abfallstoffen bestehende Torf ist ein Kohlenstoffspeicher. Aber ob solche in Mooren gebundene Torfschichten Senken oder Quellen für Treibhausgase sind, hängt von den aktuellen Bedingungen ab. Für den Schutz und die Restitution von Mooren sind die Kenntnisse dieser Zusammenhänge eine wichtige Voraussetzung.
Vom Moos zur Landschaft – Morphologie und Physiologie der Torfmoose als Voraussetzung für die Hochmoorbildung erkennen
Die mikroskopische Untersuchung von Torfmoosen lässt erkennen, welche morphologischen Voraussetzungen ihrer ausgezeichneten Wasserspeicherfähigkeit zugrunde liegen. Wasserspeicherung, kapillare Wasserleitung und durch Torfmoose bedingte Veränderung des Elektrolytgehalts lassen sich experimentell untersuchen. Aus den Ergebnissen erklärt sich die Bedeutung der Torfmoose für die Hochmoorbildung.
Abb. 21 Mikroskopische Untersuchungen an Torfmoosen lassen die morphologischen Grundlagen ihrer Wasserspeicherfähigkeit erkennen (aus Probst 1987)
Abb. 22 Wasserspeicherfähigkeit von Torfmoosen (aus Probst 1987)
Die Ionenaustauschfähigkeit von Torfmoosen kann man nachweisen, indem man die Moose Wasser mit Elektrolytgehalt aussetzt. Das zu prüfende Moospolster – etwa zwei Hand voll – wird in einem Küchensieb mehrfach mit destilliertem Wasser ausgespült und ausgedrückt, dann werden vier gewichtsgleiche Teil des Polsters zu etwa 100 g, feucht, in 3 Bechergläser mit je 200 ml unterschiedlicher Salzlösungen und einem Becherglas mit 200ml destilliertem Wasser verteilt (wie in Abb. 21 dargestellt). In jedem Ansatz wird nach 10, 20 und 40 Minuten der pH-Wert bestimmt. Die Blindprobe mit destilliertem Wasser zeigt keine Veränderung des pH-Wertes, die Probe mit der 0,01 N Calciumschloridlösung zeigt die stärkste Ansäuerung, da die Ansäuerung in gewissen Grenzen der Menge der angebotenen Kationen proportional ist und dass durch zweiwertige Calciumionen mehr H+-Ionen freigesetzt werden können als durch einwertige Kaliumionen.
Abb. 23 Versuch zur Ionenaustauschfähigkeit von Torfmoosen (aus Probst 1987)
Torfmooskultur – eine Alternative zum Torfabbau?
Zur Jahrtausendwende wurden jährlich 25 Millionen m³ Torf im Gartenbau genutzt; die auf einer Fläche von 800 km² gewonnen wurden. Wäre die gezielte Kultur und Ernte von Torfmoosen eine umweltfreundliche Alternative? Wenn man annimmt, dass damit 2500 kg Torfmoos -Trockenmasse pro Hektar und Jahr gewonnen werden könnten, würde hierzu eine Fläche von 15.000 km² benötigt, die so nicht zur Verfügung steht. Könnte die Paludikultur trotzdem ein sinnvoller und klimaschonender Zweig der Landwirtschaft werden?
Moosbeeren und Sonnentau – Nischenbildung am Extremstandort Hochmoor
Für Gefäßpflanzen sind Hochmoore ein sehr extremer Standort. Nur wenigen Arten ist es gelungen, eine ökologische Nische aufzubauen, die zu diesen Biotop passt. Der insektenfressende Rundblätterige Sonnentau und die Gewöhnliche Moosbeere, ein immergrüner, niederliegend fadenförmige wachsender Zwergstrauch, sind Beispiele für unterschiedliche Nischenbildung am selben Standort.
Schmetterlinge im Hochmoor: Hochmoor-Perlmutterfalter, Hochmoor-Gelbling und Hochmoor-Bläuling
Die drei Schmetterlingsarten sind eng an Hochmoore gebunden. Wie andere Arten gelten sie als Eiszeitrelikte, die nach der Erwärmung in den Hochmooren eine letzte Zuflucht gefunden haben. Die Raupe des Hochmoor-Perlmutterfalters ernährt sich nur von den Blättern der Moosbeere, während die beiden anderen Arten auch Heidelbeeren, Preiselbeeren und Rauschbeeren als Futterpflanzen annehmen. Die Falter sind auf nektarreiche Blüten der umgebenden Vegetation angewiesen. Die Ursachen für die Gefährdung dieser Arten werden analysiert.
Moore besitzen besondere konservierende Eigenschaften, die vor allem dem Sauerstoffmangel und dem niedrigen pH-Wert zu verdanken sind. So können in Mooren eingelagerte Werkzeuge, Waffen oder Schmuck ebenso Jahrtausende überdauer, wie Siedlungsstrukturen und Reste von Pflanzen und Tieren (und Menschen!). Dies gilt auch für Mikrostrukturen wie Pollen und Sporen, mit deren Hilfe man die nacheiszeitliche Vegetationsgeschichte rekonstruieren konnte (Pollenanalyse).
Dargie, G.C. et al. (2017): Age, extent and carbon storageof the central Congo Basin peatland complex. Nature 542, 7639, pp 1476-1487
Dierßen, K./Dierßen, B. (2008): Moore. Ökosysteme Mitteleuropas in geobotanischer Sicht. Stuttgart:Ulmer
Eigner, J. (2003): Möglichkeiten und Grenzen der Renaturierung von Hochmooren. Laufener Seminarbeiträge, 1/03, S. 23 -36, Laufen/Salzach: Bayer: Akad. f. Naturschutz u. Landschaftspflege
Ellenberg, H./Leuschner, L. (6. A., 2010): Vegetation Mitteleuropas mit den Alpen. Stuttgart: Ulmer (UTB)
Frey, W./Lösch, R. (3.A., 2010): Geobotanik. Pflanze und Vegetation in Raum und Zeit. Heidelberg: Spektrum
Garcin, Y., Schefuß, E., Dargie, G.C. et al. (2022): Hydroclimatic vulnerability of peat carbon in the central Congo Basin. Nature. https://doi.org/10.1038/s41586-022-05389-3
Gewin, V. (2020): Bringing back the bogs. Nature 578, pp. 204-208
Göttlich, K. (Hrsg.,1990) Moor- und Torfkunde. Stuttgart: Schweizerbart’sche Verlagsbuchhandlung.
Hakobyan, A., Liesack, W. (2020): Unexpectedmetabolitic versality among type II methanotrophs in the alphaproteobacteria. Biol.Chem.401(12). pp1469-1477
Hölzel, N. T. et al. (2019): Leitfaden zur Torfmoosvermehrung für Renaturierungszwecke. Deutsche Bundesstiftung Umwelt, Osnabrück.
Jäger, C. (2020): Klimaschutz braucht Moorschutz. München: Oekom
Joosten,mH:;tanneberger, F., Moen, A. (eds., 2017): Mires and peatlands of Europe.Status, distribution and conservation.Stutttgart: Schweizerbart
Kremer, B. P./Oftring,B. (2013): Im Moor und auf der Heide. Bern CH: Haupt
Kosta ,J.E. et al. (2016): The Sphagnum microbiom: new insights from an ancient plant lineage. New Phytologist 211(1), pp 57-64. doi: 10.1111/nph.13993.
LLUR (2015): Moore in Schleswig-Holstein Geschichte – Bedeutung – Schutz. Landesamt für Landwirtschaft, Umwelt und ländliche Räume des Landes Schleswig-Holstein (LLUR). 162 S
Overbeck, F. (1975): Botanisch-ökologische Moorkunde. Neumünster: Wachholtz
Page, S.E., Rieley, J.O.,Wüst, R. (2006): Lowland tropical peatland of Southeast Asia. In: Martini,I.P., MatinezCortizas, A., Chesworth. E. editors: Peatland: Evolution and records of environmental and climate changes. Chapter 7, pp 145-170
Probst, W. (1978): Zur Vegetation des Jardelunder Moores. Die Heimat 85 (Heft 10/11), S. 2 72-296
Probst, W. (1987): Biologie der Moos- und Farnpflanzen, 2. A.. Heidelberg/Wiesbaden: Quelle und Meyer
Proff, I., Furtak, S. (2022): Nasse Lawirtschaft. In: Spektrum Kompakt Feuchtgebiete, S.41-54, Heidelberg: Spekrum
Ricker, K.-M. (2021): Moore für das Klima. Die Bedeutung der Moore für den Klima- und Naturschutz kennenlernen. Biologie 5 – 10, S. 20-23, Hannover: Friedrich
Springer, P. (2017): Sphagnum als Torfersatz. GartenbauProfi, 8/13. S-48-49
Steiner, G.M. (2005): Moortypen. Stapfia 0085, S. 5-26
Succow, M. (2001): Moorkunde, 2. A., Stuttgart: Schweizerbart’sche Verlagsbuchhandlung
Succow, M., Jeschke, L. (2022): Deutschlands Moore: Ihr Schicksal in unserer Kulturlandschaft. Rangsdorf: Natur& Text
Succow, M./Joosten, H. (2001): Landschaftsökologische Moorkunde. Stuttgart: Schweizerbart’sche Verlagsbuchhandlung
Trepel, M. (2007/8): Zur Bedeutung von Mooren in der Klimadebatte. Jahresbericht des Landesamtes für Natur und Umwelt des Landes Schleswig-Holstein.
Vile, M. A. et al. (2014): N2-fixation by mmethanotrophs sustains carbon and nirtrogen accumulation in pristine peatlands. Biogeochemistry Vol121, pp 317-328, DOI:10.1007/s10533-014-0019-6
Wichtmann, W., Schröder, C. & Joosten, H. (Hrsg.) 2016: Paludikultur – Bewirtschaftung nasser Moore. Stuttgart: Schweizerbart
Für den Jahrgang 2025 ist ein Unterricht Biologie Heft mit dem Thema „Schleim in der Biologie“ geplant. Als voraussichtlicher Herausgeber dieses Heftes suche ich Autor* innen für Unterrichtsmodelle zu diesem Thema. Diese Zusammenstellung soll Interesse wecken und zur Mitarbeit ermuntern.
Schleim ist ein bisschen ekelig aber gleichzeitig auch faszinierend.
Von Ernst Haeckels Urschleimtheorie über den intelligenten Schleimozean auf Stanislaus Lems Planet Solaris, Lovecrafts Shoggothen-Schleimmonstern und der wissenschaftich begründeten Horror-Zukunftsvision des Meeresbiologen Daniel Pauly, eines Myxozäns, einer Erdepoche des Schleims, bis zum Kinderspielzeug Magic Slime, das auch Erwachsene fasziniert, zieht sich eine verzweigte Schleimspur durch die Vorstellungswelt der Menschen.
Bei Tieren und Menschen werden Schleimstoffe in Drüsen sezerniert. Sie dienen dem Schutz von Schleimhäuten und sie sind Bestandteil von Speichel und Magensaft sowie von Knorpeln, Sehnen, Haut und anderen Geweben. Für manche Tiergruppen ist die Schleimbildung besonders charakteristisch, zum Beispiel für die Mollusken. Fischen hilft ihr Schleimüberzug, den Widerstand des Wassers zu verringern und das Anheften von Schmarotzern und Krankheitserregern zu verhindern. Eine sehr primitive Fischform, die zu den Kieferlosen gehörenden Schleimaale, produzieren über ihre in der Haut gelegenen Schleimdrüsen bei Bedrohung plötzlich so viel Schleim, dass Maul und Kiemen der angreifenden Fische verstopft werden und diese den Schleimaal wieder ausspucken.
Pflanzliche Schleime findet man vor allem in Früchten und Samen (Leinsamen, Chiasamen), aber auch in Rinden, Wurzeln und Blättern (Aloe). Die stark quellfähigen Substanzen dienen der Wasseraufnahme und dem Schutz vor Austrocknung, bei Früchten und Samen auch als Klebstoff. Diese Eigenschaften machen sie auch für den Einsatz in Medizin und Kosmetik interessant. Auch Algen, Pilze und Bakterien können gute Schleimproduzenten sein (Agar). Die Schleimpilze oder Myxogastria sind eine ganz besondere Gruppe von Lebewesen zwischen Einzellern und Vielzellern, die nach ihrer häufig schleimigen Konsistenz benannt sind.
Verfolgen wir einige dieser Schleimspuren:
Physik und Chemie der Schleimstoffe
Typisch für Schleimstoffe ist ihre Quellfähigkeit und der daraus resultierender hohe Wassergehalt sowie ihre Klebrigkeit. Physikalisch gehören die Schleime zur „weichen Materie“ (soft matter, McLeish 2020), Stoffen, die sich nur bedingt dem Aggregatzustand fest oder flüssig zuordnen lassen. Das sind z. B. außer Schleimen kolloide Suspensionen (Milch), Flüssigkristalle (verwendet in Displays), Elastomere (Gummi), Tenside (Seifenschaum) oder Gele (Götterspeise). In der Strömungslehre bezeichnet man solche Substanzen auch als Nichtnewtonsches Fluide. Im Gegensatz zu Newtonschen Fluiden ändert sich ihre Viskosität, wenn sich die auf sie einwirkende Scherkräfte verändern. Damit entsprechen sie nicht dem newtonschen Elementargesetz der Zähigkeitsreibung. Dies hängt damit zusammen, dass die Scherkräfte eine Veränderung der mikroskopischen Struktur bewirken und damit die Wechselwirkungen zwischen den Teilchen des Fluids beeinflussen.
Schleime bestehen aus sehr langgestreckten Molekülfäden, die nicht einfach fließen können, wie die viel kleineren Moleküle normaler Flüssigkeiten wie Wasser oder Ethanol. Denn die Fäden verstricken sich miteinander und gehen an Kontaktpunkten Verbindungen ein. Sie können sich nur bewegen, wenn sie an den Kontaktpunkten getrennt und wieder neu zusammengefügt werden. Ein Bild dafür sind die etwas aneinander klebenden Nudelfäden auf einem Teller Spaghetti. Man kann sie entwirren und „zum fließen bringen“, wenn man sie mit einer Gabel aufwickelt und dadurch parallelisiert. In dem submikroskopischen Schleim bewirkt die Brownsche Molekularbewegung diese Entwirrung, die dafür sorgt, dass die einzelnen Molekülfäden ihren Weg durch das Gewirr finden. Die Viskosität des Schleims ist dabei außer von der Temperatur von den Eigenschaften der Fadenmoleküle abhängig. Wenn die Polymere keine einfachen Fäden sind, sondern Verzweigungen oder kammartige Strukturen besitzen, erhöht sich die Zähigkeit.
Abb. 1 Hyaluronan-Monomer
Viele tierischen Schleime bestehen aus Glykosaminoglykanen (GAG, auch als Mucopolysaccharide bezeichnet). Das sind saure Polysaccharide aus linear aneinandergereihten Disacchariden, zum Beispiel der Hyaluronsäure (Hyaluronan, von griech. Hyalos = Glas) aus dem Disaccharid aus D-Glucuronsäure und N-Acetyl-D-glucosamin (Abb.1). Mit über 50 000 Disaccharideinheiten hat Hyaloronan eine Molekülmasse von mehreren Millionen. Hyaluronan ist ein wichtiger extrazellulärer Bestandteil des Bindegewebes der Wirbeltiere. Neben der Wasserspeicherung ist ihre Druckstabilität (zum Beispiel in den Bandscheiben), und ihre Wirkung als Gelenk-Schmiermittel bedeutend. Der Glaskörper des Auges besteht zu etwa 98 % aus Wasser, Hyaluronsäure und Kollagenfasern. Im Gehirn bewirkt Hyaluronan den Wiederaufbau von Markscheiden (Remyelinisierung), weshalb ihr eine inhibitorische Wirkung bei Multipler Sklerose zugeschrieben wird. Bei Zellteilungen und Zellwanderungen scheinen Wechselwirkungen von Hyaluronsäure und Zelloberflächen eine Rolle zu spielen.
Noch größere Makromoleküle sind die Mucine, Glykoproteine mit einem zentralen Proteinfaden an den kovalent gebundenen Kohlenhydratgruppen angeheftet sind. Die Kohlenhydrate werden erst nach der Translation vor allem an die Aminosäurereste Asparagin, Serin, Threonin oder Hydroxylysin angelagert. Durch die Polysaccharide können die Mucine sehr viel Wasser binden und damit das zentrale Protein vor Abbau unter der Einwirkung von Säuren schützen. Sie spielen eine wichtige Rolle für die Barrierefunktion der Schleimhäute. (Abb. 2 ). Durch Disulfidbrücken können sich Mucinmoleküle zu noch größeren Aggregaten verbinden. An den Enden der Polysaccharidketten finden sich teilweise Sulfatgruppen oder Sialinsäuregruppen, welche den bakteriellen Abbau erschweren. Mucine werden von verschiedensten Mikroorganismen (Bakterien und Archäen, Einzeller, Pilze, Schleimpilze) und Tieren gebildet.
Andere pflanzliche Schleime und Schleime von Algen und Bakterien bestehen vorwiegend aus Polysacchariden. Dazu zählen die Pectine der pflanzlicher Zellwände, die Galactomannane aus Samen von Hülsenfrüchtlern und die quellfähigen Polysaccharide klebriger Samenschalen wie etwa bei Wegerich- Arten („Flohsamen“ des Indischen Wegerichs werden zur Regulierung der Verdauung genutzt). Auch die die Alginate und Carageene verschiedener Algen bestehen aus Polysacchariden (Abb. 3, 4 )
Abb. 3 Schleimige Jochalgen. Vor allem in Frühjahr können Jochalgen wie Spirogyra oder Mougotia (im Bild) schleimige Watten in Pfützen, Kleingewässern und Gartenteichen bilden (Foto: W. Probst)
Biofilme
Abb. 4 Bathybius Haeckelii (nach heutigen Nomenklaturregeln müsste das Epitheton klein geschrieben werden „haeckelii“) (aus Haeckel 1870)
Ernst Haeckel hat vor 150 Jahren angenommen, dass alles Leben einem Urschleim entstammen würde, der den Meeresgrund überzieht. Dieser wabbernde Glibber, so die Vorstellung, sollte ständig neues Leben hervorbringen. Thomas Henry Huxley, wie Haeckel begeisterter Anhänger von Darwins Selektionstheorie, meinte 1868 diesen Urschleim in Proben des nordatlantischen Meeresbodens gefunden zu haben und benannte die Entdeckung Bathybius Haeckelii. Haeckel war hocherfreut und schrieb in einer Publikation 1870 „ Die wichtigste Tatsache, die aus Huxley‘s sehr sorgfältigen Untersuchungen des Bathybius hervorgeht, ist, dass der Meeresgrund des offenen Ozeans in den bedeutenderen Tiefen (unterhalb 5000 Fuß) bedeckt ist mit ungeheuren Massen von freiem lebendem Protoplasma, … Dieser universelle Urschleim der älteren Naturphilosophie, der im Meer entstanden sein und der Urquell alles Lebens, das produktive Material aller Organismen sein sollte, … – er scheint durch Huxleys Entdeckungen des Bathybius zur vollen Wahrheit geworden zu sein“. (Ernst Haeckel 1870)
Abb.5 Stromatolithen in der Shark Bay an der Westküste Australiens (Foto E. Steiner 2005)
Abb. 6 Tintenstriche an Kalkfelsen der Gola Gorropu, Sardinien (Foto: W. Probst 1992)
Der von Thomas Henry Huxley beschriebene Bathybius haeckelii stellte sich allerdings schon bald als ein durch Alkoholkonservierung entstandenes anorganisches Produkt heraus (Wedlich 2019). Aber die von Mikroorganismen gebildeten und besiedelten Schleimschichten, Biofilme genannt, haben durchaus etwas Urschleimiges. Man kann davon ausgehen, dass es ähnliche Kongregationen schon seit Urzeiten gibt. Stromatolithen, Kalkstrukturen, die von schleimigen Bakterienschichten überzogen und aufgebaut werden, gelten als die ältesten Lebensgemeinschaften. Als Fossilien kennt man sie seit über 3 Milliarden Jahren und man findet sie noch heute, zum Beispiel an der Westküste Australiens (Shark Bay; Abb. 5). Für 2-3 Mrd. Jahre waren solche schleimigen Lebensgemeinschaften in den Meeren die einzigen Lebensformen. Aber ähnliche Aggregate könnten auch schon früh die Festländer besiedelt haben.
Die schleimigen Kolonien des Blaugrünen Bakteriums Nostoc, biologische Bodenkrusten und „Tintenstriche“ an Kalkfelsen sind möglicherweise solche terrestrischen bis heute überdauernde Lebengemeinschaften der Früherde (Abb. 6)., ebenso die Biokrusten, die sich auf offenen Sandbödn bilden können.
Als Biofilme bezeichnet man Schleimschichten, die eine Mischung aus Mikroorganismen (Bakterien, Archäen, Algen, Pilze, Einzeller) bestehen (Abb. 7, 8).
Abb. 7 Initialstadium eines Biofilms. Frei lebende Bakterien setzen sich fest, werfen ihre Geißeln ab und sondern Schleim ab (Grafik W. Probst)
Sie können auch einige mehrzeiligen Organismen (Rädertierchen, Fadenwürmer, Milben) enthalten, die sich von den Mikroben ernähren. Die Schleimbeläge bilden sich an Oberflächen und Grenzflächen, sowohl an Übergängen von flüssigen zu gasförmigen als auch von festen zu flüssigen Substraten. In weiterer Fassung kann man darunter auch mit Mikroorganismen angereicherte Schleimklümpchen in Flüssigkeiten verstehen. Die Schleimstoffe werden von den Lebewesen, vorwiegend von den Bakterien und Archäen, abgeschieden. Die sogenannten extrazellulären polymeren Substanzen (EPS) bestehen aus Polysacchariden, Proteinen, Lipiden und Nukleinsäuren. Sie können sehr viel Wasser binden und Hydrogele bilden, in denen Nähr- und Mineralstoffe gelöst sind. Neben verschiedenen organischen und anorganischen Partikeln können auch Gasblasen eingeschlossen werden. In einem Biofilm können in geringen Abständen sauerstoffreiche und sauerstoffarme oder -freie Bereiche liegen, die dann jeweils von aeroben bzw. anaeroben Mikroorganismen besiedelt werden. Die Oberflächen der Filme können ebenfalls sehr unterschiedlich gestaltet sein. Teilweise siedeln sich dort in das umgebende Wasser hineinragende Organismen (zum Beispiel Glockentierchen) an, teilweise bilden sich Ausbuchtungen, Poren oder Höhlen, die den Stoffaustausch erleichtern. An der Grenzschicht können Teile des Biofilms abreißen vom vorbeiströmenden Wasser weiterverbreitet werden und im Wasser Schleimflocken bilden.
Abb. 8 Entwicklung und Alterung eines Biofilms (Grafik W. Probst)
Voraussetzung für die Bildung eines Biofilmes ist, dass sich die Mikroben an einer Oberfläche festsetzen können. Dabei verändern sich die Organismen. Bei Bakterien ist es häufig mit dem Verlust der Flagellen und dem Abscheiden von Polymeren verbunden. In Biofilmen gibt es zwischen den einzelnen Mikrobenzellen einen Signalaustausch, der dafür sorgt, dass Zellteilung und Wachstum reguliert ablaufen. Dadurch wird Mangelernährung und Zusammenbruch des Systems vermieden (Quorum Sensing). Als Kommunikationsfaktor ist z. B. bei Bacillus subtilis die Abgabe von K+-Ionen nachgewiesen. Auch ein altruistischer Nährstoffaustausch von gut versorgten Mikroben zu „unterernährten“ Mikroben ist nachgewiesen.
Durch horizontalen Gentransfer können die Organismen in einem Biofilm gegenseitig ihre Genausstattung verbessern und zum Beispiel Gene weitergeben, die sie zur energetischen Nutzung bestimmter Substrate befähigen oder sie gegen Gifte resistent machen. In der Endphase der Biofilmentwicklung kommt es dann, ebenfalls durch Signalstoffe verursacht, zur Abgabe von begeißelten Formen und zur Sporenbildung. Auch abgerissene Biofilm-Flocken dienen der Ausbreitung, denn sie können sich leicht an neuen Oberflächen festsetzen und weiter wachsen.
All diese besonderen Formen der Kooperation und Vehrmehrung lassen Biofilme als Superorganismen erscheinen, in denen sich verschiedene Prokaryoten über extrazelluläre Matrices aus Makromolekülen verbinden und ihre Stoffwechselaktivitäten sehr effektiv aufeinander abstimmen können. Schon kurz nach der Entstehung des Lebens und lange vor echter Vielzelligkeit entwickelte sich so eine höhere Organisationsebene des Lebens mit echter Differenzierung der verschiedenen beteiligten Einzelzellen (vgl. Margulis 1997).
Biofilme sind sehr weit verbreitet, in allen Böden, auf Sand, auf Gesteinen auf und in Pflanzen und Tieren, in heißen Quellen und auf dem Gletschereis, in technischen Geräten, Rohren und Röhrchen, Tanks und U-Booten. Wüstenkrusten zum Beispiel sind die ersten Biozönosen lockerer Wüsten-Sandböden. Sie bestehen aus Bakterien, Algen, Pilzen und schließlich auch Flechten und Moosen. Die von den Mikroben abgegebenen Kohlenhydrate bilden nicht nur eine Matrix für die verschiedenen Lebewesen, sie verkleben auch die anorganischen Substratpartikel. Diese Krustenbildung verhindert Winderosion, fördert Wasserabsorbtion selbst aus Tau oder Nebel und führt über Luftstickstoff-Fixierung sogar zu einer Anreicherung lebenswichtiger Stickstoffverbindungen. Auch für Wattboden-Oberflächen sind Biofilme charakteristisch. Hier spielen neben Blaugrünen Bakterien Diatomeen (Kieselalgen) eine wichtige Rolle. Auch hier kommt es durch ausgeschiedene Polysaccharide zur Krustenbildung („Wattpapier“).
Besondere Biofilm-Gemeinschaften finden sich auf den Häuten und Schleimhäuten von Tieren und als Zahnbelag. Die große Bedeutung dieses Mikrobioms, das an Zellenanzahl häufig die Zellenzahl ihres Trägertieres übertrifft, wurde erst in den letzten Jahrzehnten erkannt und ist immer noch ein zentrales Forschungsthema.
Dies gilt auch für die klinische Bedeutung von Biofilmen. Trotz ihrer weiten Verbreitung wurde ihre Gefahr in der Medizin lange Zeit unterschätzt. Dabei schützen sich etwa 60 % aller mikrobiellen Krankheitserreger durch Biofilmbildung vor dem Immunsystem (Fux et al. 2005). Die Ablösung von Bakterienflocken aus Biofilmen kann zur Quelle chronisch wiederkehrender Infektionen werden, besonders bei Patienten mit geschwächtem Immunsystem. Dies betrifft zum Beispiel Krankheiten wie Blasenentzündungen, Parodontose, chronische Mittelohrentzündung oder chronische Lyme-Borreliose. Auch Biofilmbildungen auf medizinischen Instrumenten, Kathetern und chirurgischen Implantaten können der Ausgangspunkt von Infektionen sein. In sehr vielen chronischen Wunden lassen sich Biofilme nachweisen.
An technischen Konstruktionen aus Metall können Biofilme Korrosion hervorrufen. Auch Luftbefeuchter und Verdunstungskühlanlagen sind besonders anfällig für Biofilmbildungen.
Die schleimigen Mikrobengemeinschaften können aber auch sinnvoll genutzt werden, zum Beispiel in der biologischen Abwasserreinigung oder bei der mikrobiellen Laugung von Erzen (selektive Anreicherung bestimmter Mineralien). Selbst Bodenschadstoffe wie Mineralölrückstände können durch entsprechende Mikroorganismen-Gemeinchaften abgebaut werden.
Eine wichtige Nutzanwendung sind die Biofilme aus lebenden Essigbakterien zur Herstellung von Essig aus Ethanol (Essigmutter) oder das schleimige Konglomerat aus Essigbakterien und Hefepilzen („Teepilz“), mit dem sich gesüßter Schwarztee in den Kombuchatrank (Teekwaß) umwandeln lässt.
Kombucha – das Geheimnis eines Zaubertrankes
Abb. 9 Teepilz (Kombucha) – aus einem ebay-Angebot von ilja.g94 (11.2021)
„Wenn sie nach einem Mittel mit fast magischen Kräften suchen, das Sie stimulieren und verjüngen kann, so gibt es keine Möglichkeit, die Sie näher an Ihr Ziel bringt, als der Kombuchapilz-Tee. Wir machen Ihnen diese Versprechungen, und wenn Sie nicht hundertprozentig befriedigt von dem herrlich schmeckenden Tee und dem von ihm bewirkten gesundheitlichen Segnungen sind, werden wir Ihnen alle Auslagen einschließlich der Transportkosten zurück erstatten!“
Mit diesem einleitenden Satz wird von einem Anbieter von Kombucha-Tee im Internet geworben.
Der Glaube an Wunder wirkende „Allheilmittel“ ist wahrscheinlich so alt wie die Menschheit. Als eine solche Wunderdroge wird der Tee“pilz“ oder Kombucha immer wieder benannt und angeboten. Ähnlich wie bei Kefirknollen handelt es sich dabei um eine enge Gemeinschaft von Bakterien und Hefepilzen. In diesem Falle gewinnen Sie Ihre Lebensenergie vor allem aus dem Abbau des im gesüßten Tee reichlich enthaltenen Rohrzuckers. Dabei werden auch andere Inhaltsstoffe der Teeflüssigkeit genutzt und im Stoffwechsel verarbeitet. Als Endprodukte entstehen nicht nur Kohlenstoffdioxid und verschiedene Karbonsäuren (vor allem Essigsäure, Milchsäure, Ethanol,Gluconsäure und Glucuronsäure), im Kombuchatrank konnten auch Aminosäuren, Usninsäure, die Vitamine B1, B2, B3, B6, B12, Folsäure sowie Vitamin C nachgewiesen werden.
Der sogenannte Pilz ist in biologischem Sinne natürlich kein Pilz. Er besteht aus einer gallertigen Masse aus Polysacchariden, vorwiegend Zellulose, die als Matrix für verschiedene Hefearten und Bakterien dient (Abb. 11 ). Dieses schleimige Aggregat kann Tochteraggregate bilden oder durch Teilung vermehrt werden. Ähnliche Mikrobengemeinschaften sind verhältnismäßig weit verbreitet, ja, sie dürften eine ganz charakteristische Lebensweise von Mikroorganismen, insbesondere von Prokaryoten, Hefen und Schimmelpilzen darstellen. Für die menschliche Ernährung genutzt, werden sie z.B. auch als „Essigmutter“, als Kefir oder als Wasserkefir.
Die „Teepilz-Symbiose“ ist zunächst ein farbloses, fast völlig transparentes Schleimklümpchen. Bei Temperaturen zwischen 12 und 30 °C (Vorzugstemperatur: 23-27 °) und genügendem Sauerstoffzutritt wächst ein solches Klümpchen in mit Zucker gesüßtem Tee zu einem weißlich durchscheinenden Gebilde unregelmäßiger Form mit schleimigem Äußeren heran. Bei weiterem Wachstum flacht es sich ab und nimmt schließlich die ganze Oberfläche des Gefäßes ein. Wenn die gesamte Flüssigkeitsoberfläche vom „Teepilz“ bedeckt ist, wächst der hellgrau durchschimmernde Schirm verstärkt in die dritte Dimension und die Bezeichnung Bio“film“ passt dann eigentlich nicht mehr. Aus dem scheibenförmigen Gebildet wachsen lamellenähnliche Strukturen nach unten in die Kulturflüssigkeit. Mit der Zeit sinkt das ganze Aggregat tiefer in die Flüssigkeit ein. Unter Beibehaltung der schleimigen Oberfläche nimmt es dabei eine immer dunklere, bräunlich-hellgraue Farbe an und wird außerordentlich zäh. Wenn die äußeren Bedingungen ungünstig werden, wenn z.B. in der Kulturlösung kein Zucker mehr enthalten ist, kann das Aggregat seine „Zusammenarbeit“ beenden und sich auflösen. Ein solcher Vorgang kann mit dem Sterben eines vielzelligen Organismus verglichen werden (vgl. Margulis 1997). Die einzelnen Mikroorganismen – Hefezellen und Bakterienzellen – müssen dabei nicht absterben. Isoliert sind sie jedoch nicht zu den gleichen Stoffwechselleistungen in der Lage wie im Verband. Wohl aber können sie unter günstigen Umweltbedingungen wieder zu einem neuen Verband zusammen treten. So gesehen ist eine Analogie zu den Keimzellen vielzelliger Organismen gegeben.
Abb. 10 Teepilz – Mikroskopische Aufnahme eines Schleimklümpchens mit Bakterien- und Hefezellen, ca. 400x (Foto W.Probst 2002)
Im Gegensatz etwa zur Flechtensymbiose, bei der es sich bei jeder „Flechtenart“ um eine ganz dezidierte Kombination einer oder weniger bestimmter Pilz- und Algenarten handelt, ist die Zusammensetzung des Kombuchaaggregates variabel. Auch das Kulturmedium hat Einfluss auf diese Zusammensetzung und damit auch auf die abgegebenen Stoffwechselprodukte und die Inhaltsstoffe des Kombucha-Getränkes. Mit dem Kombuchatrank nimmt man jedoch immer neben den organischen Säuren, unter denen vor allem die Gluconsäure und die Glucuronsäure sich förderlich auf die Entgiftungsprozesse in der Leber auswirken sollen, auch lebende Hefe- und Bakterienzellen auf. Soweit sie verdaut werden, können daraus Vitamine z.B. der B-Gruppe freigesetzt werden. Wenn sie lebend in den Dünndarm gelangen, können sie sich günstig auf die Zusammensetzung und Wirkung der Darmflora auswirken. Gesundheitsfördernde und heilende Wirkungen sind damit vor allem im Zusammenhang mit einem starken Glauben durchaus möglich. Schädliche Wirkungen jedenfalls braucht man bei dem Genuss von Kombucha nicht zu befürchten, soweit das Getränk mäßig eingenommen wird. Bei einer mäßigen Dosierung dürfte der geringe Alkoholgehalt auch für Kinder nicht schädlich sein. Dasselbe gilt für den Säuregehalt. Im Gegenteil: Man kann sagen, dass das regelmäßige Vorhandensein von Essigsäurebakterien dafür sorgt, dass durch den niederen pH-Wert gefährliche Mikroben in dem Kombuchaaggregat kaum Fuß fassen können.
Der Kombuchatrank soll in China schon vor 2000 Jahren bekannt gewesen sein. Den Namen Kombucha führt man auf die japanische Bezeichnung für Tee „Cha“ und für einen ebenfalls für Teezubereitung, Salate und Gemüse verwendeten Tang „Conbu“(Laminaria japonica) zurück. Außer in Ostasien hat der Teepilz und das mit seiner Hilfe gewonnene Gärgetränk auch in Russland eine lange Tradition. Dies schlägt sich in Namen wie Teekwass, Kargasok-Teepilz, Fungojapon, Chinesischer Teepilz, Japanischer Teepilz , Mandschurischer Pilz, Russische Blume, Russische Qualle, oder Wolgameduse nieder. Die sagenhaften Heilwirkungen des Getränkes werden durch Namen wie Heldenpilz, Champignon de la Charité oder Champignon de Longue Vie beschrieben.
Wissenschaftliche Untersuchungen des „Teepilzes“ begannen erst im 20. Jahrhundert. 1913 beschrieb Lindau in den Berichten der Deutschen Botanischen Gesellschaft über eine Teepilzkultur, die er von einem Herrn Dr. Gisevius aus dem Kurland erhalten hatte (Medusomyces gisevii Lindau). Bei der mikroskopischen Untersuchung konnte Lindau nur Hefepilze erkennen und so beschrieb er das Gebilde als „Medusomyces Gisevii nov. spec. et nov.gen.“ Noch im selben Jahr konnte der Botanikprofessor Lindner nachweisen, dass der Teepilz keine eigene Art ist, sondern vielmehr aus einem Konglomerat verschiedener Bakterien- und Hefe-Arten gebildet wird. Als bakteriellen Hauptbestandteil isolierte er das Schleim-Essigbakterium Acetobacter xylinum und der wichtigste pilzliche Organismus konnte von ihm als der auch aus dem afrikanischen Hirsebier („Pombe“) bekannte Hefepilz Schizosaccharomyces pombe bestimmt werden. Weitere Hefepilze der Gattungen Torula, Torulopsis, Pichia, Candida, Saccharomycodes und Mycoderma konnte nachgewiesen werden. In den Jahren nach dem ersten Weltkrieg brachten aus Russland zurückkehrende Kriegsgefangene Teepilzkulturen mit. Die Herstellung des Teekwass wurde auch in Deutschland bekannt. In den 20er und 30er Jahren wurden erste wissenschaftliche Untersuchungen über die Stoffwechselleistungen der Teepilz-Symbiose durchgeführt. Dabei konnten im Teekwass Milchsäure, Essigsäure, Ethanol und auch Gluconsäure nachgewiesen werden. In einer neueren Untersuchung stellte Reiß ( 1987) die quantitative Zusammensetzung der Teepilz-Gärungsprodukte fest. Er konnte auch zeigen, dass das verwendete Substrat (Schwarztee bzw. Lindenblüten-, Pfefferminztee, Colagetränk und Bier) Einfluss auf die Zusammensetzung der Gärungsprodukte hat.
Unter natürlichen Bedingungen dürften ähnliche Mikrobenaggregate vermutlich überall dort vorkommen, wo zuckerhaltige Pflanzensäfte in größeren Mengen auftreten. Vor allem dürfte dies der Blutungssaft von Bäumen und anderen verletzten Gewächsen sein, eventuell auch Blüten mit reichlicher Nektarproduktion oder süße, große Früchte.
In der Literatur wird beschrieben, dass „Teepilze“ auch technisch genutzt wurden (Lindner 1917/ 1918 und Lakowitz 1928 nach Meixner 1983). Man ließ die Aggregate meterdick und zentnerschwer werden und nutzte sie zur Herstellung von Handschuhleder und für Gasballonhüllen.
Sukkulente
Eine mögliche Anpassung von Pflanzen an trockene Standorte ist die Sukkulenz: Blätter oder Sprossachsen, seltener auch Wurzeln, können stark verdickt sein und ein sehr wasserhaltiges Gewebe aus großlumigen Zellen enthalten. Der Zellsaft weist meist eine schleimige Konsistenz auf, die vor allem von Polysaccharideden aus Glucose, Mannose, Galactose und Xylose sowie Aminosäuren und Carbonsäuren zustande kommt. Das Gel dient nicht nur der Wasserspeicherung sondern auch einer besonderen Form der Photosynthese, der CAM-Photosynthese (von Crassulacean Acid Metabolismus). Diese sukkulenten Pflanzen öffnen nachts ihre Spaltöffnungen und speichern CO2 in organischen Säuren, zum Beispiel Äpfelsäure. Tagsüber bleiben die Spalten geschlossen und das CO2 aus den Carbonsäuren wird freigesetzt und für die Photosynthese verwendet. Dadurch kann der Wasserverlust durch Transpiration stark eingeschränkt werden.
Abb. 11 Aloe aristata-Blatt mit schleimigem Inhalt (Foto W. Probst 2021)
Sukkulente kommen in vielen verschiedenen Pflanzenfamilien vor. Besonders bekannt sind Kakteen und Kakteen-ähnliche Euphorbien oder die Dickblattgewächse (Crassulaceae), deren Name schon auf ihr Blattsukkulenz hinweist. Auch bei den Lilienverwandten gibt es viele Blattsukkulente, zum Beispiel die Agaven und die Aloe-Arten. Besonders berühmt für ihren Blattschleim ist die Echte Aloe (Aloe vera). Die Pflanze wird seit dem Altertum medizinisch genutzt und gilt bis heute als medizinisches Wundermittel. Im Internet finden sich zahlreiche Angebote, die das Pflanzengel nicht nur zur äußerlichen Anwendung gegen Verbrennungen, Sonnenbrand, Mückenstiche, Ekzeme, Geschwüre, unreine Haut und Entzündungen aller Art empfehlen, sondern auch zur inneren Anwendung bei Husten, Sodbrennen, Diabetes, Allergien und Reizmagen bzw. Reizdarm. Entsprechend groß ist die Palette der Aloe-vera-Produkte.
Abb. 12 Aloe-Schleim (Foto W.Probst 2021)
Wie bei allen Wundermitteln und Allheilmittel ist auch hier Vorsicht geboten. Medizinische eindeutige Nachweise für die vielseitigen Heilwirkung gibt es nicht. Eine Verwendung des Gels bei zu trockener Haut kann sicherlich nicht schaden und zumindest die kühlende, Juckreiz stillende Wirkung bei Insektenstichen gilt auch als gesichert. Bei der innerlichen Anwendung ist Vorsicht geboten, denn der in den äußeren Blattgeweben enthaltene gelbliche Saft enthält Aloin, ein Stoff aus der Gruppe der 1,8-Dihydrohxyanthracene. Der bitter schmeckende Stoff ist möglicherweise krebserregend außerdem stark abführend.
In alten Streuobstwiesen kann man Apfelbäume finden, die im Winter so grün sind wie Immergrüne. Auch Pappeln und Birken können sehr dicht mit Misteln (Viscum album) besetzt sein. Diese immergrünen Pflanzen wachsen nicht nur auf sondern auch in ihren Wirtsbäumen. Sie treiben ihre Wurzeln bis in das Holz ihrer Wirte und zapfen deren Wasserleitungsbahnen an. So gewinnen sie Wasser und Mineralstoffe, für die Produktion von Kohlenhydraten sorgt ihr eigener Fotosyntheseapparat in den grünen Blättern (Abb. 13).
Abb. 13 Misteln (Viscum album) auf Schwarz-Pappeln, Eriskircher Ried, 19.4.2014 (Foto W. Probst)
Misteln sind zweihäusig, d. h., eine Mistelpflanze trägt immer nur männliche oder weibliche Blüten. Allerdings können Misteln auch auf Misteln parasitieren. Wenn Parasit und Wirt unterschiedlichen Geschlechts sind, entsteht so der Eindruck einer einhäusigen Pflanze.
Abb. 14 Mistelpflanze (Viscum album) an Birke. Dank ihrer streng dichotomen Verzweigungen haben ältere Mistelpflanzen eine nahezu perfekte Kugelform (Foto W. Probst, 2016)
Geringer Mistelbesatz schadet einem Baum wenig. Aber sehr viele Misteln können dazu führen, dass der Wirtsbaum – vor allem bei niedrigen Temperaturen im Winter, wenn die Wasseraufnahme eingeschränkt ist – vertrocknet. Das ist dann allerdings auch das Ende seiner grünen Parasiten, aber diese haben sich in der Zwischenzeit schon stark vermehrt. Denn die Misteln haben eine sehr effektive Form der Samenverbreitung. In ihren weißen Beerenfrüchten ist normalerweise ein Same enthalten. Er ist eingebettet in einen zähen Schleim (Abb. 16). Die Früchte reifen im Spätherbst und bleiben bis zum Frühjahr, manchmal sogar bis zum Frühsommer erhalten. Die Beeren werden sehr gerne von Drosseln, vor allem von den danach benannten Mistel-Drosseln, gefressen. Die Samen werden aber nicht verdaut. Sie werden von den Drosseln wieder ausgeschieden, und auch ein Teil des zähen Schleims überlebt die Darmpassage. Solche ausgeschiedenen Mistelsamen mit Schleimfäden kann man bei genauer Beobachtung nicht selten an Ästen von Bäumen finden. Dieser biologische Zusammenhang war schon den Römern bekannt. Außerdem waren Drosseln schon im alten Rom eine beliebte Delikatesse und sie wurden mit Vogelleim gefangen, den man aus Mistelbeeren herstellte (meist allerdings aus der Eichenmistel Loranthus europaeus, die in Deutschland nur an wenigen Stellen vorkommt. Daher das römische Sprichwort).
Abb. 15 Mistelfrucht (Foto W. Probst 2016)
Kleinere Vögel wie Meisen fressen nur den äußeren, weniger klebrigen Teil des Fruchtfleischs und streifen den Samen mit seinem klebrigen Mantel an den Zweigen ab. In beiden Fällen werden die Samen so durch die Vögel an Zweige und Äste von Bäumen geklebt.
Im Übrigen war die Mistel schon in der Antike eine verehrte Heilpflanze, nicht nur bei Griechen und Römern, sondern auch bei Kelten (für Asterix-Fans: ein wichtiger Bestandteil des Zaubertrankes des Druiden Mirakulix). Die Verehrung spiegelt sich bis heute in dem vor allem in angelsächsischen Ländern beheimateten Brauch wieder, grüne Mistelzweige zu Weihnachten ins Zimmer zu hängen. Dass man sich unter solchen in Wohnungen aufgehängten Mistelzweigen küsst bzw. küssen darf, gehört in England und den Vereinigten Staaten zu den Weihnachtsbräuchen und hat seinen Ursprung vermutlich in heidnischer Vorzeit. In der modernen Heilkunde spielen vor allem die Mistellektine als mögliche Tumorhemmer und Stimulatoren des Immunsystems eine gewisse Rolle. Lektine sind Proteine oder Glykoproteine, die sich spezifisch mit Zellmembranen verbinden können und von dort biochemische Reaktionen auslösen.
Abb. 16 Der Schleim in der Mistelfrucht (Fotos W. Probst)
Früchte oder Samen, die klebrigen Schleim für ihre Verbreitung nutzen, kennt man auch von anderen Pflanzen. Ein gutes Beispiel sind die Früchte der Wegerich Pflanzen. Die meisten Pflanzen mögen es nicht so gerne, wenn man auf ihnen herum trampelt, nicht so – wie schon der Name sagt – die Wegeriche. Sie gedeihen gerade an solchen Stellen, an denen Konkurrenten durch Vertritt ausgeschaltet werden. Besonders trifft dies in unserer heimischen Flora auf den Breit-Wegerich zu. Man findet ihn fast nur auf Wegen oder anderen häufig begangenen und befahrenen Stellen.
Abb. 17 Gequollene Samen des Breit-Wegerichs (Plantago major) in wässriger Methylenblau-Lösung. Die Schleimhüllen nehmen die Farblöung nicht so schnell auf. (Foto W. Probst 2010)
Ab Juli kann man seine Früchte finden. Es sind Kapseln, die sich an langen Ähren entwickeln. Wenn sie reif sind, löst sich ein Deckel ab und die Samen werden ausgestreut. Die Samen sind von einer Klebschicht umgeben, die allerdings erst klebt wenn sie nass wird – ähnlich wie der Kleber auf der Rückseite einer Briefmarke. In diesem Zustand hat sie ausgezeichnet an Hufen aber auch an Schuhen. Das ist ein Grund dafür, dass der Breit-Wegerich heute auf der ganzen Erde anzutreffen ist. Nach der Besiedelung durch weiße Siedler kam die Pflanze auch nach Nordamerika. Wo die Siedler mit ihren Planwagen und Tieren ihre breiten Spuren hinterlassen hatten, breitete sich die vertrittfeste Pflanze aus und weitere Siedlerkarawanen schadeten ihr nicht. Den Indianern galt diese Pflanze bald als Zeichen der Kolonisten und sie nannten sie „Fußspur des weißen Mannes“ (Abb. 18).
Abb. 18 Breit-Wegerich in Pflasterfuge (Foto W. Probst 2019)
Schleimige Samen werden vom Menschen traditionell in Heilkunde und Medizin genutzt. „Flohsamen“ werden aus den Samenschalen der Wegerich Arten Plantago indica und Plantago afra hergestellt und dienen wegen ihrer wasserbindenden Eigenschaften der Verdauungsregulation. Sie werden sowohl bei Verstopfung als auch bei Durchfall eingesetzt. Leinsamen sind ein altbekanntes Mittel gegen Verstopfung. Eine ganz hippe neue Schleimspeise sind Chia-Samen (von der mexikanischen Salvia hispanica), die zum Beispiel als Brotzusatz oder in Müsli-und Joghurtspeisen Leinsamen mittlerweile an Popularität übertreffen. Auch die traditionelle Krankenmahlzeit „Schleimsuppe“ aus Haferflocken erhält ihre schleimige Konsistenz aus den Schleimstoffen der zu Flocken gewalzten Haferkörner.
Schneckenschleim
Abb. 19 Spanische Wegschnecken (Arion vulgaris) bei der Paarung (Foto W. Probst 9.2000)
Schneckenschleim ist zäh und klebrig. Besonders haftstark ist der Schleim der Spanischen Wegschnecke. Das erfährt man, wenn man ein solches Schneckentier mit der Hand von Pflanzen absammelt. Der Reiz stimuliert die Nacktschnecke zur Schleimproduktion. Dieser Schleim ist nicht nur besonders zäh, er enthält auch Bitterstoffe. Natürliche Schneckenfresser wie Kröten und Igel halten sich deshalb bei Spanischen Wegschneckten zurück. Gartenbesitzern sind diese Schnecken ein Graus, denn sie können besonders in feuchten Sommern in solchen Massen auftreten, dass man kaum eine Chance hat, ein Gemüsebeet anzulegen, da alle Setzlinge oder Keimlinge sofort abgefressen werden. Salatpflanzen, Zucchini und Kohlrabi werden ebenso vertilgt wie Basilikum und Rucola oder Zierpflanzen, vor allem Lilienverwandte.
Mit ihrer Raspelzunge können die Schnecken nicht nur Pflanzenblätter im Nu klein raspeln, sie vertilgen alle möglichen Arten von organischem Abfall und sie schrecken auch vor Kannibalismus nicht zurück. Um ihre Pflanzen zu schützen, gehen Gartenbesitzer mit Messern, Scheren, Salz oder Fallen auf Schneckenjagd. Das giftige Schneckenkorn wirkt anziehend auf Schnecken und führt bei Verzehr zu einem schleimigen Tod. Da der enthaltene Wirkstoff Methiocarp ein gefährliches Nervengift ist, dass nicht nur alle Arten von Schnecken tötet, sondern auch für andere Tiere und für den Menschen gefährlich werden kann, wurde seine Zulassung als Molluskizid in der EU im Oktober 2013 und generell 2019 zurückgenommen.
Die reizabhängige Schleimproduktion der Spanischen Wegschnecken wird seit einiger Zeit genutzt um medizinische Produkte zu testen, die für Schleimhäute – zum Beispiel bei Augenuntersuchungen – eingesetzt werden sollen. Die Wegschnecke dient als Tiermodell, um chemische Substanzen auf ihre Verträglichkeit zu untersuchen. Je mehr Schleim abgegeben wird, desto stärker ist der Reiz.
Daran kann man schon erkennen, dass der Schleim der Schnecken eine Schutzfunktion hat. Er hält Fressfeinde ab, aber er enthält auch bakterizide und fungizide Wirkstoffe. Für die kriechende Fortbewegung hat er eine wichtige Funktion, indem er als Gleit-und Haftmittel wirkt: Im Ruhezustand ist das Schleimgel ziemlich fest und klebrig. Wirken Scherkräfte ein, geht es in einen flüssigeren, gleitfähigen Zustand über. Dadurch kann die Schnecke durch die Muskelbewegungen die physikalischen Eigenschaften dieses Schleims nach ihrem Bedarf variieren. Die wellenförmig von vorne nach hinten über die Unterseite laufenden Muskelkontraktionen ermöglichen eine gleitende Fortbewegung. Allerdings bleibt dabei immer eine gewisse Schleimmenge am Untergrund haften, weshalb ständig Schleim nachproduziert werden muss. Das bedeutet einen ziemlichen Material- und Energieaufwand und vor allem einen großen Flüssigkeitsbedarf: Wird es zu trocken, „geht den Schnecken der Schleim aus“.
Wichtiger Bestandteil des Schneckenschleims sind die Faserproteine Kollagen und Elastin. Kollagen besteht aus drei umeinander gewundenen Polypeptidketten und kommt bei allen Stämmen der vielzelligen Tiere vor, Elastin bildet ein Netzwerk aus Proteinfäden die über die Aminosäure in Lysin vernetzt sind. Im Gegensatz zu Kollagen ist Elastin dehnbar. Beide Stoffe kommen auch im Bindegewebe der Wirbeltiere vor und sie sind für eine straffe, faltenfreie Haut verantwortlich. Die äußerliche und innerliche Anwendung von Schneckenschleim als Schönheits- und Anti-Aging-Mittel hat deshalb Konjunktur.
Spitzen-Schleimer: Die Schleimaale
Abb. 19 Atlantischer Schleimaal (Myxine glutinosa) (Grafik W. Probst 2021)
„Es wimmelten dort in krausen Gemisch, zu hässlichen Klumpen geballt“ die Schleimaale, so könnte man mit einer Verszeile aus Schillers Taucher die Lebensumstände der atlantischen Neunaugenverwandten Myxine glutinosa beschreiben. Die fischähnlichen Lebewesen mit aalförmigen Körper aus der Gruppe der Kieferlosen (Agnatha) haben ihr Erscheinungsbild in den letzten 300 Millionen Jahren kaum verändert. So gesehen sind sie ein Erfolgsmodell der Evolution.
Lange Zeit galten Schleimaale oder Inger – es sind rund 80 Arten beschrieben – als Schädlinge, vor allem für die Grundnetzfischerei. Sie fressen die gefangenen Fische an und machen den Fang dadurch unbrauchbar. Andererseits spielen sie im Ökosystem der Tiefsee eine wichtige Rolle bei der Aufbereitung von Abfallstoffen, vor allem großer Kadaver. Sie wurden deshalb als „Totengräber der Tiefsee“ bezeichnet. Wirtschaftliche Bedeutung hat seit einiger Zeit die Haut der Schleimaale, da sie zur Lederherstellung verwendet („Aalleder“) werden kann. In Ostasien werden Schleimaale auch als Speisefisch genutzt. Vor allem im Westen der USA wird Aalleder in größerer Menge produziert. Die Schleimaale werden von LKWs zu den Lederproduktionsstätten gefahren. So kam es wohl 2017 in Oregon zu einem Verkehrsunfall mit einem solchen Schleimaal-Transporter, bei dem 3,4 t dieser Tiere auf die Straße gerieten und alles mit ihrem Schleim überzogen. Von ProSieben wurde ein Video dieses Unfalls in der Hoffnung auf Werbeeinnahmen ins Netz gestellt , auch in National Geographic wurde der Unfall beschrieben.
Walkadaver in der Tiefsee mit Schleimaalen (Grafik W. Probst 2021)
Der Schleimaal-Schleim ist ein ganz besonderer Glibber und deshalb ist er in den Fokus der Wissenschaft geraten. Einmalig ist seine extrem hohe Wasserspeicherfähigkeit. In gequollenem Zustand bestehen nur 0,004 Gewichts% des Schleims aus Proteinen und Polysacchariden, der Rest ist Wasser. Eine weitere Besonderheit ist, dass er nicht nur Mucine sondern 15-30 cm lange Proteinfäden enthält, die Ähnlichkeiten mit der Spinnenseide aufweisen und wie diese eine extrem hohe Reißfestigkeit besitzen. Produziert werden diese beiden Schleimbestandteile in speziellen Drüsen in zwei verschiedenen Zelltypen, die entweder den Proteinfaden oder das Mucin produzieren. Über Poren werden diese beiden Bestandteile gleichzeitig ins Wasser ausgestoßen, mit dem sie sofort interagieren und zu gewaltigen Schleimmassen aufquellen. Potenzielle Anwendungsgebiete wären zum Beispiel neue, biologisch abbaubare Polymere, Gele als Füllmaterial und Mittel, um Blutungen bei Unfallopfern und Chirurgie-Patienten zu stoppen. Die Hoffnung, den Schleim exakt nachbilden zu können, sind bis jetzt allerdings gering, da seine genaue Zusammensetzung und Bildung sehr komplex erscheinen. Aber eventuell könnte das Prinzip nachgebildet werden und dann Stoffe ergeben, die ähnliche Eigenschaften haben.
Eine weitere Besonderheit des Schleimaals ist erwähnenswert: Nachdem er sich durch Schleimausstoß seine Gegner vom Hals gehalten hat, besteht für ihn nun das Problem, seinen eigenen Schleim wieder loszuwerden. Dabei nützt ihm seine große Beweglichkeit und Biegsamkeit: Er kann in seinen Schlangenleib einen Knoten machen, diesen lässt er dann langsam vom Schwanz bis zum Kopf wandern und streift damit den Schleim ab.
Schleimhäute
Abb. 21 Aufbau der menschlichen Mundschleimhaut (Grafik W. Probst 2021)
In der Medizin werden als „Schleimhäute“ die Schutzschichten bezeichnet, die innere Organe auskleiden, zum Beispiel die Verdauungsorgane, die Atmungsorgane und die Geschlechtsorgane. Im Gegensatz zur äußeren Haut besitzen Schleimhäute fast keine Hornschicht und keine Haare dafür meist Schleimdrüsen oder einzelne Zellen, die Schleim abgeben also Mucine produzieren. Sie sind aufgebaut aus einer ein-oder mehrzelligen Epithelzellenschicht, einer Bindegewebsschicht und teilweise auch noch einer Muskelschicht. Die Epithelzellen können zur Oberflächenvergrößerung kleine Ausstülpungen (Mikrovilli) tragen oder auch mit Cilien besetzt sein (Wimpernepithel). Durch die enge Verbindung mit dem Lymphsystem haben Schleimhäute eine wichtige Schutzfunktion gegen eindringende Krankheitserreger.
Der Mundspeichel wird in unterschiedlicher Zähigkeit von kleinen Speicheldrüsen in der Mundschleimhaut und von den großen Speicheldrüsen Ohrspeicheldrüse, Unterzungendrüse und Unterkieferdrüse gebildet, bei einem erwachsenen Menschen am Tag im Durchschnitt etwa ½ L. Die schleimigen Bestandteile sind Mucine, außerdem enthält der Mundspeichel das Kohlenhydrate-Verdauungsenzym Ptyalin, Ca-,Na- K- und Cl-Ionen. Antikörper (Immunoglobulin A), Laktoferin, Lysenzym und Histatin Er wirkt antiseptisch und wundheilend, weshalb es sehr wirkungsvoll ist, seine Wunden zu lecken.
Aber auch Krankheitserreger können durch Speichel übertragen werden. Deshalb ist das Auf-den-Boden-Spucken zu vermeiden und in manchen Ländern verboten, anderenorts aber auch durchaus verbreitet, zum Beispiel in China und in Indien. Früher war das Spucken auch in Mitteleuropa üblich, auf Bahnhöfen wurden Spucknäpfe aufgestellt und in Zugabteilen fand man das Schild „Nicht auf dem Boden spucken!“
Das Ausspucken ist in Indien und Südostasien oft mit dem Kauen von Betel verbunden. Für einen als“Pan“ bezeichneten Betelbissen werden die kleingehackten Arekanüsse der Betelpalme (Areca catechu) mit Löschkalk in ein Blatt des Betelpfeffers (Piper betle) gewickelt, zur Abmilderung des bitteren Geschmacks werden meist einige Gewürze wie Pfefferminze oder Lakritze zugefügt. Die Mischung wird etwa eine Viertelstunde gekaut. Das führt zu einer starken Anregung des Speichelflusses und zu einer intensiven Rotfärbung des Speichels durch die in der Arekanuss enthaltenen Phlobatannine. Überflüssige Flüssigkeit wird ausgespuckt – auf dem Boden und an Wänden und Mauern kann man überall die roten Flecken erkennen. Die Wirkung der enthaltenen Alkaloide ist so ähnlich wie die von starkem Kaffee.
Ein verwandter Brauch ist das Kauen von Tabak. Beim Schnupftabak werden dagegen die Nasenschleimhäute zur Aufnahme des Nikotin genutzt. Das durch den Schnupftabak ausgelöste Niesen ist ein Teil des Genusses und ein großes Schnupftuch gehört dazu. Früher waren solche Stofftaschentücher auch für das Schneuzen bei normalem Erkältungsschnupfen sehr üblich und ein beliebtes Verlegenheitsgeschenk bei Geburtstagen. Erst durch den Siegeszug der Papiertücher sind die Stofftaschentücher zur Aufnahme des Nasenschleims weitgehend verschwunden.
Ein genetisch bedingte Schleimkrankheit ist die Mukoviszodose oder Cystische Fibrose (CF). Bei den Schleim absondernden Zellen funktionieren Chloridkanäle in der Zellmembran nicht mehr richtig, da in dem für das Zellkanal-Protein zuständigen Gen eine Mutation aufgetreten ist. Dadurch wird der osmotisch bedingte Wasseraustritt von den umgebenden Zellen in den abgesonderten Schleim gebremst und der Schleim bekommt eine sehr zähe Konsistenz. Dies betrifft nicht nur Nasenschleimhäute und Bronchien sondern auch Sekrete der Bauchspeicheldrüse, der Leber (Gallen), der inneren Geschlechtsorgane, der Speiseröhre, des Darms und der Schweißdrüsen. Die Folge sind Funktionsstörungen unterschiedlichster Art. Die Symptome der Krankheit zeigen sich bereits in der frühen Kindheit und die Krankheit ist unheilbar und führt meist über kurz oder lang zum Tode.
Bei Menschen liegt das mutierte Gen auf dem langen Arm von Chromosom 7. Bisher sind über 2000 verschiedene Mutationen dieses Gens bekannt, das auch CFTR-Gen genannt wird (Cystic Fibrosis Transmembran Conductance Regulator bezeichnet das transmembrane Kanalprotein). Die häufigste Mutation ist eine Punktmutation, die zu einem fehlen der Aminosäure Phenylalanin an Position 508 des CFTR-Proteins führt.
In Europa kommt auf 2000 Geburten ein Fall von Mukoviszidose. Die Krankheit wird autosomal rezessiv vererbt. Dank verbesserter Therapiemöglichkeiten hat sich die Überlebensrate von Mukoviszidose-Patienten in den letzten Jahrzehnten deutlich verbessert. Aus evolutionsbiologischer Sicht ist es verwunderlich, dass sich die krankheitserregenden Allele dieses Gens sich in so hoher Konzentration in der Population erhalten haben. Das kann man sich eigentlich nur durch einen Selektionsvorteil des heterozygoten Genotyps erklären. Möglicherweise besteht bei den Heterozygoten eine höhere Resistenz gegen Tuberkulose.
Neben der symptomatischen Behandlung wird seit einiger Zeit auch versucht, ein gesundes Gen in die Zellen einzubauen, dass dann für die Produktion eines funktionsfähigen CFTR-Proteins fungiert (Gentherapie). Bisher gibt es bei diesen Versuchen aber noch keine funktionierenden Ergebnisse. Eine weitere Möglichkeit wäre eine Behandlung mit entsprechender mRNA, die dann direkt an Ribosomen in die richtigen Proteine translatiert werden könnte. Auch hier liegen bisher (2021) noch keine positiven Ergebnisse vor.
Zum Schluss
Unser Weg auf biologischen Schleimspuren ist nun erst einmal zuende, aber natürlich wäre noch vieles zu entdecken, zum Beispiel
die schleimigen Netze der Myxogastria, die sich in Labyrinthen zurecht finden und die effektivsten Verbindungen zu verschiedenen Nahrungsquellen finden, und die deshalb sogar menschlichen Netzwerkplanern als Vorbild dienen;
der Schleim der Amphibien, der ihre fast unverhornte Haut vor Austrocknung schützt aber auch giftige Substanzen zur Feindabwehr einhalten kann und ihre Eier mit einer glibberigen Hülle umgibt;
die Bedeutung von Schleimstoffen bei der Fortpflanzung;
die verschiedenen Schleimstoffe in unseren Speisen, von Haferschleimsuppe und Chiamüsli, Remoulade und Hering in Aspik bis zu glibberigem Kaviar und schleimigen Austern.
Vielleicht haben Sie Lust auf weitere Entdeckungen? Die Internetslinks eröffnen einige Möglichkeiten.
Abb. 22 Grasfrosch mit Laich, 11.3.2017 (Foto W. Probst)
Quellen
Buslau, S.-J., Hembd, C. (1999): Kombucha. Das Gesundheitselexier aus China. München: Heyne
Flemming, H.-C. (2000):Biofilme – das Leben am Rande der Wasserphase. In: Nachrichten aus der Chemie 48 (4): 442 f, 2000
Flemming, H.-C., Wingender, J. (2001): Biofilme – die bevorzugte Lebensform der Bakterien. Biologie in unserer Zeit 31 (3): 169
Flemming, HC., Wingender, J., Szewzyk, U. et al. Biofilms: an emergent form of bacterial life. Nat Rev Microbiol14, 563–575 (2016). https://doi.org/10.1038/nrmicro.2016.94
Lange, O. L. Hrsg.(2013): Biological Soil Crusts: Structure, Function, and Management. Springer
Lem, S: Solaris 1961, Deutsch von I. Zimmermann-Göllheim. Berlin:Ullstein 2006
Lüttig, A., Kasten, J. (2003): Hagebutte & Co. Blüten, Früchte und Ausbreitung europäischer Pflanzen. Not drei Nahrungen Allweil mangelnden RES Januar zwei ja tuln: Fauna-Verlag
Margulis, L.( 1997): From Kefir to death. In: Margulis, L./Sagan, D.(1997): Slanted truths., New York: Copenicus-Springer, 83-90
Margulis, L., Sagan, D. (1986): Microcosmos. Four billion years of microbial evolution. Berkeley…:University of California Press
Mayser, P. u.a.: The yeast spectrum of the tea fungus Kombucha. Mycoses 38: 289-295, 1995
McLeish, T. (2020): Soft matter. A very short introduction. Oxford Univ. Press
Meixner, A. 1983(1):1-4, 1984(2): 32-34, 1985(1): 7-10: Combucha, der Teepilz. Südwestdeutsche Pilzrundschau, Stuttgart
Nelson, D. L., Cox, M. M. (2009): Lehninger Biochemie. 4.A. Berlin/Heidelberg: Springer
Treffpunkt: 10.00h am Wanderparkplatz im Brochenzeller Wald an der Landstraße L 329 zwischen Ettenkirch und Brochenzell, von Oberteuringen kommend links, kurz vor dem Kreisverkehr am Ortseingang von Brochenzell
Auf diesem Spaziergang gehen wir eine kleine Runde durch dieses schöne Waldgebiet mit abwechslungsreichem Baumbestand. Besonders interessieren uns die Pflanzen an den feuchten Wegrändern und Gräben.
Dauer: Etwa 2,5 Stunden
Ersatztermin bei schlechtem Wetter: 21.08.2015
Wälder am Rand des Schussenbeckens
Größere zusammenhängende Waldgebiete sind im heutigen Oberschwaben ziemlich selten. Im Laufe der mehrtausendjährigen Siedlungsgeschichte ist die typische kleinräumige Landschaft aus Felder – heute vielfach Obstplantagen -, Wiesen und kleinen Waldstücken entstanden. Das größte Waldgebiet, der Altdorfer Wald etwa zwischen Vogt und Wolpertswende gelegen, hat immerhin eine Längsausdehnung von ca. 17 km. Dagegen ist das Waldgebiet , das sich am westlichen Rand des Schussenbeckens etwa von Ravensburg bis Meckenbeuren erstreckt, mit knapp 8 km deutlich kleiner. Die geplante Umgehungsstraße für Meckenbeuren könnte es noch weiter verkleinern. Trotzdem kann man in diesem Wald stundenlang wandern. Mehrere Bäche entwässern das Gebiet zur Schussen hin. Sie haben sich zum Teil ziemlich tief in die Jungmöräne eingeschnitten.Trotzdem ist der Wald ein guter Puffer, der einen Teil des Regenwassers speichert und dadurch vor Überschwemungen schützt.
SIG Sigmaringen, EHI Ehingen, UL Ulm, SLG Saulgau, BC Biberach, STO Stockach, ÜB Überlingen, KN Konstanz, TT Tettnang, RV Ravensburg, WG Wangen, MM Memmingen (aus Köhler, A.: Vom Wesen und Werden der oberschwäbischen Landschaft. In Ott, St. (Hrsg.), 1971: Oberschwaben. Otto Maier Verlag Ravensburg)
Mit dem Bodenseebecken wird das untere Schussental im Allgemeinen zum Landschaftsraum Bodensee-Schussen-Becken zusammengefasst, der am Ende der Würmeiszeit durch den Rheingletscher geprägt wurde.
Zum Exkusionsverlauf
Exkursionsweg
Wegbegleitende Hochstaudenflur
Wegränder sind häufig die artenreichsten Biotope eines Waldgebietes. Wenn sie nicht zu oft gemäht werden, entwickelt sich hier vor allem hochwüchsige, nährmineralliebende Stauden, aber auch Sträucher und Lianen. Besonders im Spätsommer und Herbst sind ihre Blüten ein wichtiges Futter für viele Nektar oder Pollen fressende Insekten. Auf unserem Spazierweg fallen besonders die großen Blüten- und Fruchtstände des Wiesen-Bärenklaus auf – wir messen beim höchsten 2,50m. Sie werden ebenso wie die der Wald-Engelwurz von Bienen, Hummeln, Schwebefliegen, Käfern und Schmetterlingen besucht.
Auch der Wasserdost liefert reichlich Nektar. Wegen der etwas tieferen Blütenkronröhren ist er vor allen für Schmetterlinge attraktiv. Wie Riesen-Schachtelhalm, Gilbweiderich und Kohl-Kratzdistel ist er Zeichen für einen feuchten Standort. Quellhorizonte sind typisch für die das Schussenbecken umrandenden Jungmoränen, in die immer wieder Lehm- und Tonschichten eingelagert sind. Neben dem befestigten Waldweg wurden zwei Gräben angelegt, um das Wasser der Quellhorizonte aufzunehmen.
Zitronenfalter (Gonepteryx rhamni) auf Kohl-Kratzdistel (Cirsium oleraceum) (Foto M. Pohl, 14.8.2021)
Der Name „Kohl“-Kratzdistel weist darauf hin, dass die Pflanzen als Gemüse genutzt werden können. Besonders die großen, weichen Grundblätter können wie Spinat zubereitet werden. Der Wurzelstock enthält reichlich Inulin und kann zu Mehl verarbeitet werden, das zum Andicken von Speisen dient. Auch der gegarte Blütenboden kann ähnlich wie Artischockenblütenstände genutzt werden. Kohl-Kratzdisteln werden in Japan angebaut. Ein weiterer Korbbühler, der Gewöhnliche Rainkohl, kann als Jungpflanze ebenfalls für Gemüse und Salate verwendet werden. Die einjährige, selten zweijährige Pflanze ist schon verblüht und überall stehen die weitgehend abgestorbenen Fruchtstände.
Ein auffälliges, häufiges Gras der Wegränder ist der Riesen-Schwingel mit langen überhängenden Rispen und großen begrannten Ährchen. Er ist typisch für feuchte Laubwälder und wächst besonders häufig in Auwäldern entlang der Wasserläufe. Zwei andere Gräser, die erst jetzt, für Gräser also ausgesprochen spät blühen, sind das Pfeifengras und die Rasen-Schmiele, die beide auf feuchten Standort hindeuten. Beim Pfeifengras ist der lange Halm knotenlos, alle Knoten finden sich an der Halmbasis. Pfeifengrashalme wurden deshalb früher zum reinigen langer Pfeifenrohre verwendet. In Norddeutschland heißt das Gras „Benthalm“. Dieser Name weist auf die Nutzung der Halme zum Aufbinden hin. Besenried wird das Gras genannt, weil man aus den Halmen früher auch Besen angefertigt hat. Pfeifengraswiesen sind typisch für teilweise trockenfallende Moore, auch entwässerte Hochmoore.
Die Rasen-Schmiele hat große, stark aufgeteilte Blütenrispen mit sehr kleinen Ährchen und sehr raue, gerillte Blätter, die im Durchlicht auffällig gestreift erscheinen. Die durchscheinenden Rillen sind die Stellen, über die sich die Blätter bei Trockenheit einrollen können. Außer auf feuchten bis nassen Wiesen, Weiden und Sümpfen gedeiht sie in feuchten, lichten Laubwäldern und in Quellfluren.
Nach der waldökologischen Standortkartierung Baden-Württemberg ist der typische Wald des Schussenbeckens und seiner Randmöränen ein Buchenwald mit Tanne, Edellaubbäumen – also Ahorn-Arten, Ulmen, Kirschen, Linden und Eschen – Eichen und Hainbuchen. Die ebenfalls relativ häufigen Rot-Fichten und Wald-Kiefern sind vor allem auf Aufforstungsmaßnahmen zurückzuführen.
In den Bereichen in denen der Weg durch die Kronen der Laubbäume ziemlich stark beschattet ist, treten die Hochstauden am Wegrand zurück. Vor allem die dichten Kronen von einigen Winter-Linden (gekennzeichnet durch braune Haare in den Winkel der Blattadern auf der Unterseite) beschatten die Wegränder. Hexenkraut und Stinkender Storchschnabel sind häufig, in den moosreichen Nadelwaldbeständen gedeiht Wald-Sauerklee. An einigen Stellen haben sich große Bestände des einjährigen Großen Springkrautes (mpatiens noli-tangere) entwickelt, das im Gegensatz zu dem eingeschleppten Drüsigen Springkraut typisch für sehr schattige Standorte ist.
Winter-Linde und Weiß-Tanne (Foto W. Probst,20.8.2021)
Hier wachsen nebeneinander Rot-Fichte und Weiß-Tanne und wir beschäftigen uns mit den Unterschieden dieser beiden einheimischen Nadelgehölze („Fichte sticht, Tanne nicht“) und auch mit den Gattungsunterschieden von Tanne und Fichte: Für die Gattung Fichte sind die braunen Nadelstielchen charakteristisch, die nach dem abfallen der Nadeln am Zweig stehen bleiben. Für die Gattung Tanne ist die verbreiterte grüne Nadelbasis charakteristisch, die nach dem abfallen eine glatte Zweigoberfläche zurücklässt.
Gelbbauchunken (Bombina variegata)
Gelbbauchunke (Bombina vriegata) aus dem Brochenzeller Wald (Foto Lilli Schiller, 14.8.2021)
An der tiefsten Stelle des Weges, der die L 329 mit der Kreisstraße 7731 von Meckenbeuren nach Taldorf verbindet, liegen wir in einen kleineren Weg nach rechts ab. Er wird von einem Wasser führenden Graben begleitet, in dem große Bestände der Wasser-Schwertlilie stehen. In einer Wegpfütze entdecken wir Kaulquappen der Gelbbauchunke und nach genauem hinschauen sowohl in der Pfütze als auch im angrenzenden Graben die Unken selbst.
Kaulquappen der Gelbbauchunke in Wegpfütze, 20.8.2021 (Foto W. Probst)
„Eine Gefährdung für die Vorkommen der Gelbbauchunke geht vor allem von Lebensraumverlusten aus, wie sie beispielsweise durch das Verfüllen von Tümpeln, durch Ausbau von unbefestigten Waldwegen und Rückegassen, aber auch durch natürliche Sukzession (vor allem die Verbuschung) der Pionierbiotope auftreten. Infolge einer räumlichen Verinselung, also einer Fragmentierung der Habitate (beispielsweise durch Straßen oder intensive Landwirtschaft), werden Vorkommen voneinander getrennt. Dadurch werden der genetische Austausch zwischen den Populationen und die Zuwanderung von außen eingeschränkt, was letztlich die Überlebensfähigkeit der isolierten Bestände gefährdet“.(Wikipedia)
Die Unken sind nach der Roten Liste der Bundesrepublik Deutschland stark gefährdet (2). Sie sind sowohl nach dem Bundesnaturschutzgesetz als auch nach der Fauna-Flora-Habitat-Richtlinie streng zu schützen. Innerhalb Deutschlands sind sie als „Verantwortungsart“ eingestuft.
Gelbbauchunke in einer Wegpfütze im Brochenzeller Wald (Foto W. Probst, 20.8.2021)
Die Unkenpfütze hat Verbindungen zum wegbegleitenden Graben. Dort gedeihen Hänge-Segge (Carex pendula), Gewöhnlcher Blutweiderich (Lytrum salicaria), Gewöhnliches Mädesüß (Filipendula ulmaria), Kohl-Kratzdistel (Cirsium oleraceum), Sumpf-kratzdistel (Cirsium palustre) und Pfeifengras (Molinia caerulea).
Unkenbiotop im Brochenzeller Wald 26.8.2021 (Foto W. Probst)
auf dieser Seite findet ihr alle wichtigen Informationen zum Exkursionsangebot im Rahmen der Veranstaltung „Regionale außerschulische Lernorte Oberschwabens“.
In der folgenden Übersicht haben wir die geplanten Exkursionsorte und -zeiten angegeben. Sollten einige Teilnehmer*innen auf öffentliche Verkehrsmittel angewiesen sein, müssen wir die Exkursionsziele eventuell ändern. Dies werden wir auf der ersten Veranstaltung in Oberteuringen klären.
Ausrüstung: Außer einer exkursionsgerechten Kleidung
Schreibzeug und Notizbuch
Mobiltelefon mit Fotofunktion,
Botanikerlupe und Fernglas
Behälter (Stofftasche oder Plastikbeutel; kleine Sammelbehälter ) zum Unterbringen von Sammlungsstücken
eventuell eine Sitzunterlage für Pausen im Gelände
Studienleistungen: Portfolio zu den Exkursionen, in dem zu jeder Exkursion mindestens zwei Objekte, Themen, Aktivitäten … beschrieben und kommentiert werden. Bei den Objekten können auch Scans oder Fotos verwendet werden, sodass auch ein digitales Portfolio möglich ist.
Artenkenntnis : Wichtiges Ziel der Exkursionen ist die Verbesserung der Artenkenntnis. Deshalb werden auf jeder Exkursion von zwei Teilnehmenden alle angesprochenen Arten notiert. Sie werden anschließend auf dieser Homepage veröffentlicht.
Kommentare und Fragen sind erwünscht, während den Exkursionen, als Kommentar auf dieser Seite oder per E-Mail. Wenn sie Pflanzen- oder Tierarten betreffen, ist ein beigefügtes Foto hilfreich.
Die erste Veranstaltung findet am 8. Mai in Oberteuringen statt. Von Weingarten/Ravensburg aus ist der Ort über die B 33 (Richtung Meersburg) zu erreichen. Außerdem gibt es eine Busverbindung von Ravensburg nach Oberteuringen, die ihr unter folgender URL finden könnt:
Auf die Exkursionen freuen sich Sabrina Brendle und Wilfried Probst.
Übersicht über die Exkursionsorte und -termine
Exkursionsorte, geändert am 8.5.2021
Zeit
Treffpunkt
Themen
8.5.21 13.30-17.30h
Oberteuringen, Franz-Roth-Platz
Baumgeschichten; Bestimmungsschlüssel; Suchen und Finden; Kletterpflanzen; Wiese
15.5.21 10.00-14.00
Weingarten, Freibad
Natur wahrnehmen und erleben
29.5.21 13.30-17.30h
Oberteuringen, NSG Altweiherwiese*
Landschaftsgeschichte; Gräser; Zeigerwerte von Pflanzen; Saumbiotope; Biber
12.6.21 10.00-1400h
Völlkofen, Grillhütte
Wildkräuter und ihre Verwendung
3.7.21 13.30-17.30h
Wilhelmsdorf, Naturschutzzentrum
Insekten, Landschaftsgeschichte:Hoch- und Niedermoor
24.7.21 18.30-22.30h
Wilhelmsdorf, Naturschutzzentrum
Fledermäuse
* geändert am 8.5.2021
8. Mai 2021 Oberteuringen
Anfahrt zum Treffpunkt in Oberteuringen, Franz-Roth-Platz
1. Von Ravensburg auf der B 33 Richtung Meersburg bis Oberteuringen, Ortsteil Hefigkofen und weiter bis Ortsteil Neuhaus.
2. In Neuhaus an der ersten Kreuzung links in die Teuringer Straße abbiegen.
3. Der Straße bis zum Ortsschild „Oberteuringen“und bis zur 30iger Zone folgen, dann erste Abzweigung links zum Parkplatz „Franz-Roth-Platz“ abbiegen.
Vorlagen: Google maps
Oberteuringen
Foto W.Probst
Oberteuringen ist ein Beispiel für die lange Besiedelungsgeschichte Oberschwabens und des Bodenseegebietes. Die älteste Urkunde, welche die Existenz der Siedlung Teuringen belegt, stammt aus dem Jahre 752. Dabei handelt es sich um einen Beleg für die Schenkung Teuringens und einiger anderer Siedlungen an das Kloster St. Gallen. Eine solche lange Geschichte ist durchaus charakteristisch für Ortschaften deren Namen mit – ingen endet. Dabei handelt es sich um alemannische Siedlungen die vermutlich im fünften und sechsten Jahrhundert gegründet wurden und mit denen die intensivere landwirtschaftliche Bewirtschaftung Oberschwabens begann. Diese lange Siedlungsgeschichte war landschaftsprägend. Rund um die vielen kleinen Ortschaften, Weiler und Einzelhöfe ist eine reich strukturierte Landschaft entstanden. Große zusammenhängende Waldgebiete sind selten, auch die landwirtschaftlichen Flächen sind meist kleiner strukturiert. Erst die Industrialisierung in jüngerer und jüngster Zeit hat viele Ortschaften durch Gewerbegebiete und neue Wohngebiete sehr anwachsen lassen. Das gilt auch für Oberteuringen, das mittlerweile über 5000 Einwohner zählt und gerade neue Bebauungspläne aufstellt.
Ein Grund dafür, dass der erste außerschulische Lernort unserer Exkursionen ein solcher aus biologischer Sicht eher durchschnittlich erscheinender Ort Oberschwabens ist und nicht ein spektakuläres Naturschutzgebiet wie der Federsee oder das Wurzacher Ried, liegt daran, dass wir zeigen wollen, dass ergiebige außerschulische Lernorte eigentlich überall zu finden sind, ein zweiter, dass ich seit 14 Jahren in Oberteuringen wohne.
Literatur: Sanktjohanser, G. K. -Hrsg. (2002): Obeteuringen – Ein Streifzug durch die Jahrhunderte. Gemeinde Oberteuringen
Rotach bei Oberteuringen (Foto W. Probst)
Oberteuringen liegt an der Rotach, etwa in der Mitte ihres knapp 40 km langen Laufes über etwa 225 Höhenmeter vom Pfrunger-Burgweiler Ried bis zum Bodensee bei Friedrichhafen. Nach Argen und Schussen ist die Rotach der dritte größere Zulauf in den nordöstlichen Bodensee. Dieser kleine Fluss macht in Oberteuringen einen durchaus naturnahen Eindruck, der aber vor allem Renaturierungsmaßnahmen zu verdanken ist. Ursprünglich standen an der Rotach 22 Mahl- und Sägemühlen, die Oberteuringer Mühle beherbergt mittlerweile das Teuringer Kulturzentrum. Das Betreiben dieser Mühlen erforderte viele wasserbauliche Maßnahmen mit Stauwehren und Kanälen, die den natürlichen Bachlauf stark veränderten.
Gefälle der Rotach (aus Wikipedia)
Besondere Bäume am Sankt Martinsplatz
Der heutige Kirchenbau Sankt Martinus stammt aus der Zeit 1516/1517, der Ort war aber schon lange vorher der Standplatz einer christlichen Kirche. 1846 erhielt der Kirchturm seine heutige weithin sichtbare, über 60 m hohe Spitze. Der Platz zwischen Kirche und Rathaus, der Sankt Martinsplatz, ist das Zentrum Oberteuringens.
Unser Interesse galt vier besonderen Baumarten am Sankt Martinsplatz. Jeweils eine Gruppe beschäftigte sich kurze Zeit mit einer der Arten und versuchte, sie in einer möglichst kurzen Form eindeutig zu charakterisieren. Zwei „ahnungslose“ hielten sich während dessen außer Sichtweite auf. Dann versammelten sich alle vor dem Rathaus und die Gruppen charakterisierten ihren Baum. Alle vier Bäume wurden – es war zugegebenermaßen nicht sehr schwierig – sofort erkannt.
Gewöhnlicher Trompetenbaum (Catalpa bignonioides)
Trompetenbaum am 8.5.2021, Foto A.WinterTrompeebaum im Juni, Foto W. Probst
Weitere Namen: Zigarrenbaum, Bohnenbaum; der Name ist aus der Sprache der Cherokee übernommen und bedeutet „Bohnenbaum“.
Heimat: Südosten der Vereinigten Staaten, vor allem in Auwäldern und an Flussufern, 1726 durch den englischen Naturforscher Mark Catesby (1683-1749) von Carolina nach Europa gebracht.
Der Baum kann 15-18 m hoch werden. Alte Trompetenbäume bilden Absenkeräste, die einwurzeln (auf der Insel Mainau zu sehen!) und damit der Ausbreitung dienen.
Die herzförmigen bis schwach gelappten, glattrandig Blätter treiben erst sehr spät aus. Beim Zerreiben riechen sie unangenehm. In letzter Zeit kommt es in Deutschland zur Auswilderung, was vermutlich mit der Klimaerwärmung zusammenhängt.
Das Verbreitungsgebiet von Catalpa deckt sich etwa mit dem ursprünglichen Siedlungsgebiet Chirokee und vier weiterer Indianerstämme, die wegen ihrer Anpassung an die Lebensweise der Kolonisten auch als die „ Fünf zivilisierten Nationen“ bezeichnet wurden. Sequoyah (1763-1843), Sohn einer Cherokee-Indianerin und eines europäischen Händlers erfand die Cherokee Schrift, die heute noch für die Cherokee-Sprache verwendet wird. Der Mammutbaum Sequoia sempervirens wurde nach ihm benannt. Der Vertrag von New Echota von 1835 führte zur Vertreibung der Cherokee aus dem südöstlichen Waldland in Carolina und Georgia in ein karges Territorium im Staate Oklahoma. Bei dieser gewaltsamen Umsiedlung, die als „Trail of Tears“ in die Geschichte eingegangen ist, kamen vermutlich um die 8000 der Deportierten ums Leben (teilweise nach Wikipedia).
Purpur-Magnolie (Magnolia liliiflora)
Purpur-Magnolie – Magnolia liliiflora, 8.5.2021,Foto A. Winter
Heimat: China (Prov. Yunnan und Hubei); als Zierbaum in China weit verbreitet.
Das Gehölz ist meistens fast von der Basis an verzweigt und erreicht etwa 5m Wuchshöhe. Die Krone ist meist breit, Stamm und Äste sind oft unregelmäßig gekrümmt. Die Zweige sind hellgrau bis braun und nicht behaart. Auch an dickeren Stämmen bleibt die graue Rinde glatt.
Die Magnolien (Magnolia) sind eine Pflanzengattung der Familie der Magnoliengewächse mit über 200 Arten in Ostasien und Nordamerika. Ihren Namen gab ihr Linné zu Ehren des französischen Botanikers Pierre Magnol (1638–1715). Einige Magnolien haben sind beliebte Ziergehölze vor allem die Stern-Magnolie und die Tulpen-Magnolie, ein Hybrid aus Magnolia denudata und Magnolia liliiflora.
Magnolien sind sehr ursprüngliche Blütenpflanzen. Bei einer auf genetischen Analysen beruhenden Rekonstruktion einer Urblüte kam eine Blüte heraus, die dem Habitus der heutigen Magnolienblüten sehr ähnlich sieht.
Rekonstruktion einer Urblüte nach Sauquet, H. et al. (2017): The ancestral flower of angiosperms and its early diversification.Nature communications, DOI: 10.1038/ncomms16047
Gewöhnlicher Judasbaum (Cercis siliquastrum)
Gewöhnlicher Judasbaum – Cercis siliquastrum am Martinsplatz in Oberteuringen, 8.5.2021, Foto A. Winter
Heimat: Südeuropa bis Vorderasien
Angeblich hat sich Judas an einem solchen Baum erhängt. Andere Namen: Salatbaum, Liebesbaum, Stammhülsenbaum
Die Gattung hat einen Verbreitungsschwerpunkt in China (5 Arten), 4 Arten kommen in Nordamerika und eine in Zentralasien vor.
Typisch ist die Kauliflorie (Stammblütigkeit). Die biologische Erklärung ist, dass dies auch schwerere Tiere wie Kleinsäugern und Vögeln die Bestäubung ermöglicht. Kauliflore Pflanzen gibt es fast nur bei tropischen Pflanzen.
Gemeindemitteilungen Oberteuringen,20.11.20
Aus unserer Gemeinde
Der von Michaela und Manuel Knöpfler gespendete Cercis reniformis (= C. canadensis*) -Baum, auch Ju-dasbaum genannt, verschönert ab sofort den Vorplatz der Kirche St. Martin und soll als Zeichen des Lebens gerade in diesen Zeiten Mut machen. Auch wenn der Baum ausgerechnet den Namen des Jün-gers trägt der Jesus verraten und an die Römer ausgeliefert hat, nahm Jesus Schicksal durch ihn seinen Lauf. Mit seiner Auferstehung finden wir heute Hoffnung im Glauben auf das ewige Leben. Der Baum steht zwischen 2 Bänken, die bereits im Jahr 2011 ebenfalls von Michaela und Manuel Knöpfler gespendet wurden. Diese laden zum Innehalten ein und ermöglichen den Blick auf die wunderschöne Kirche St. Martin.
*Die Blattform spricht dafür, dass es sich bei der Art um Cercis siliquastrum handelt (W. Probst)
In dem gemulchten Beet, in dem der Judasbaum steht, entdeckten wir den Fruchtkörper einer Spitz-Morchel (Morchella elata).
Spitz-Morchel (Morchella elata) unter dem Judasbaum neben der Martinskirche in Oberteuingen; man beachte die Feuerwanze(!), 9.5.2021 (Foto S. Probst)
Der Unterschied zu den Echten Zypressen (Cupessus) besteht darin, dass Scheinzypressen stärker abgeflachte Zweige und zweierlei schuppenartige Blätter sowie kleinere, kugelige Zapfen besitzen und Samen früher reifen. Die ebenfalls sehr ähnlichen Lebensbäume (Thuja) haben im Gegensatz zu den Scheinzypressen kleine, längliche Zapfen. Die etwa fünf Arten (Chamaecyparis) sind in den nördlicheren Breiten Nordamerikas und Ostasiens verbreitet. Die Oregon-Scheinzypresse kann in ihrer Heimat bis 65 m hoch werden – so hoch wie der Kirchturm von St. Martin!
Das hellgelbe, harzfreie Holz wird für Schiffsbau und Möbel verwendet. In Europa ist der Baum, von dem es zahlreiche Sorten gibt, ein häufiges Ziergehölz. Mittlerweile gibt es wild wachsende Vorkommen.
Artenarmut mitteleuropäischer Wälder im Vergleich mit Nordamerika und Ostasien
In Mitteleuropa gedeihen viele Gehölzarten aus Nordameria oder dem nördlichen Ostasien, die dort unter ähnlichen Klimabedingungen wie hier existieren können. Dies hängt damit zusammen, dass die Waldvegetation Mitteleuropas während der vor etwa 2,6 Millionen Jahren beginnenden Kaltzeiten fast vollständig vernichtet wurde. Im Gegensatz zu Ostasien und Nordamerika, wo die Hauptgebirgsketten vorwiegend von Norden nach Süden verlaufen,war der Vegetation Mitteleuropas beim Vordringen der kaltzeitlichen Gletscher ein Rückzug nach Süden durch die Alpenkette weitgehend versperrt. Darin sieht man den Grund dafür, dass die mitteleuropäische Gehölzvegetation sehr viel artenärmer ist, als die entsprechenden Pflanzengesellschaften in Nordamerika und Ostasien. Im Pliozän, vor dem Beginn der Kaltzeiten (des Pleistozäns) kamen viele der heute bei uns angepflanzten Arten oder nahe Verwandte dieser Pflanzen auch in Mitteleuropa vor. Dies ist ein Argument mancher Forstleute, nun in Mitteleuropa die Aufforstung mit amerikanischen und asiatischen Baumarten zu versuchen, die mit dem Klimawandel besser zurechtkommen könnten.
Gehölze an der Rotach
Im Ortsgebiet von Oberteuringen wird die Rotach von einer Vielfalt einheimischer Gehölze gesäumt. Wir lernten sie kennen, indem wir für sie einen Bestimmungsschlüssel bastelten.
Diese Blattmerkmale werden auf einem weißen Laken ausgelegt.
Die Aufgabe besteht nun zunächst darin, zu jedem der sechs hier grün markierten Endpunkte des Bestimmungsganges Beispiele zu finden. Alle Blätter, die zu einer Art gehören, werden auf einem Haufen angeordnet. Anhand der gefundenen Blätter werden mögliche Merkmalsalternativen zur weiteren Bestimmung besprochen. Alle Arten werden mit Namen beschriftet.
Fotos A. Winter
Seifenkraut (Saponaria offcinalis)
Am Wegrandentdecken wir einen großen Bestand des Seifenkrautes. Das Nelkengewächs wird von einer Teilnehmerin, die sich sehr gut auskennt, entdeckt, obwohl die zart rosavioletten Blüten noch lange nicht entwickelt sind. Der Name der Pflanze weist auf ihre frühere Verwendung als Seifenersatz hin. Alle Pflanzenteile insbesondere die Wurzelstöcke enthalten Wasseroberflächen-entspannende Triterpensaponine. Wir zerreiben einige Triebe und schütteln Sie in einem Behälter mit Wasser und wir können die Schaumbildung beobachten.
Seifenkraut-Extrakte werden bis heute bei der schonenden Reinigung von alten Textilien und Möbelstücken verwendet (Wikipedia).
Foto A. Winter
Suchen und finden
Alle Teilnehmenden erhalten eine Suchkarte für eine Pflanzenart:
Die Arten werden ziemlich schnell gefunden: Bär-Lauch (Allium ursinum), Winter-Schachtelhalm (Equisetum hiemale), Einbeere (Paris quadrifola), Schuppenwurz (Lathraea squamaria). Letztere ist ein völlig Chlorophyll-freier Parasit an Laubbäumen.
Schuppenwurz – Lathraea squamaria – an der Rotach bei Oberteuringen, Foto W. Probst
Kletterpflanzen
Je höher eine Pflanze wächst, desto kräftiger muss ihr Stamm sein. Aber das gilt nicht für alle! Kletterpflanzen nutzen die Stabilität ihrer Unterlagen.
Bei der Brücke über die Rotach wachsen zwei Kletterpflanzen-Arten, die verholzte Liane Gewöhnliche Waldrebe (Clematis vitalba) und der krautige, jedes Jahr neu aus dem unterirdischen Wurzelstock auswachsende Echte Hopfen (Humulus lupulus). Die Waldrebe hält sich mit ihren rankenden Blattstielen an der Unterlage fest, der Hopfen windet mit seiner Sprossachse um die Unterlage, und zwar so, dass in der Seitenansicht ein S zu erkennen ist (Rechtswinder). Diese Richtung des Winden ist bei Kletterpflanzen im allgemeinen genetisch festgelegt, d. h. die Pflanzen können nur nach rechts oder nach links winden. Bei Linkswindern erkennt man in der Seitenansicht ein Z.
Lianen können hoch in Bäume hinaufklettern und viel Laubwerk entwickeln, dabei bleiben ihre Sprossachsen viel dünner als die Stämme der Bäume. Sie müssen aber fast gleich viel Wasser transportieren. Deshalb ist es wichtig, dass ihre Wasserleitungsbahnen sehr effektiv sind. Lianen haben deshalb die weitesten Tracheen aller Pflanzen (Durchmesser bis 0,7 mm). Auch die Leitungsbahnen der Waldrebe kann man schon mit bloßem Auge sehen. Wir schneiden einen etwa 1,50 m langen Sprossabschnitt der Waldrebe heraus: es gelingt ohne Mühe, durch diesen Stab Luft in ein Wasserglas zu blasen. Bei der Erweiterung der Leitungsbahnen gibt es allerdings eine Grenze: Werden die Durchmesser zu groß, reichen die Adhäsion und Kohäsionskräfte der Wassermoleküle nicht mehr aus um den hydrostatischen Unterdruck auszugleichen. Es bilden sich Luftblasen und die Wassersäule reist ab („Gasembolie“).
Spross der Gewöhnlichen Waldrebe- Clematis vitalba – quer- mit großlumigen Tracheen, Foto A. Winter
Wiesen
Wiesen und Weiden sind in unserem Klima fast ausschließlich Folgen landwirtschaftlicher Nutzung. Sie können sich nur halten, wenn sie regelmäßig von Weidetieren abgefressen oder gemäht werden. Aber sie machen mittlerweile in Mitteleuropa 50 % der landwirtschaftlich genutzten Fläche und 20 % der Gesamtfläche aus. Die Art der Bewirtschaftung ist für die Biodiversität entscheidend.
Typisch für Wiesen ist ihre Schichtung. Ähnlich wie die Frühjahrsblüher im Wald so haben die Wiesenpflanzen der Unterschicht ihre beste Entwicklungsmöglichkeiten im Frühjahr, wenn die Wiese noch nicht hoch gewachsen sind.
Wir sortieren die Wiesenpflanzen nach der Zugehörigkeit zur Unterschicht, Mittelschicht und Oberschicht.
Schichten einer Wiese
Die ersten Landlebewesen
Kolonie von Nostoc commune
Auf dem Weg zurück zum Ortszentrum bzw. zum Franz-Roth-Platz kommen wir an eine vertetene ziemlich feuchte Wegstelle mit eigenartig schwärzlichen Belägen, die mit Wasser zu olivfarbenen Gallertklumpen aufquellen. Es handelt sich um das Blaugrüne Bakterium Nostoc. So ähnlich könnten die ersten Lebewesen ausgesehen haben, die vor mehr als 3 Milliarden Jahren das Festland besiedelten.
Wasseramsel (Cinclus cinclus) – haben wir auf der Exkursion zwar nicht gesehen, kann hier aber regelmäßig beobachtet werden. Immer wieder hörten wir den Gesang von Amsel, Buchfink, Zilp-Zalp (Weidenlaubsänger) und Mönchsgrasmücke
Kletterpflanzen
Echter Hopfen (Humulus lupulus) (S-Winder)
Gewöhnliche Waldrebe (Clematis vitalba) (Z-Winder; außerdem dienen die Blattstiele und Spindeln zwischen der Fiedern als Ranken)
„Wenn wir eine Gesellschaft schaffen wollen, die die Natur wirklich liebt und ihr mit Ehrfurcht begegnet, müssen wir den Mitgliedern dieser Gesellschaft Erlebnisse in der Natur anbieten, die ihr Leben verändern.“ Joseph Cornell
Längst ist bekannt, dass das reine Wissen über die Bedrohung und die Schutzwürdigkeit der Natur allein nicht ausreicht. Aus diesem Grund stand die Exkursion am 15. Mai 2021 unter dem Thema „Natur wahrnehmen und erleben“.
Innerhalb der 4 Stunden mussten die Studierende der Pädagogischen Hochschule alle ihre Sinne einsetzten. Der Tag begann zunächst mit kleineren Kennenlernspielen um wach zu werden und die eigene Konzentration hochzufahren.
Hören
Der erste Sinn, das Hören, wurde als erstes angesprochen. Die Studierende sollten bei der Aktion „Geräusche hören“ sich auf ihr Gehör verlassen und die Geräusche in ihrer Umwelt wahrnehmen. Schritt für Schritt wurden sie an die Aktion herangeführt. Zunächst durften sie einfach darauf los hören, danach wurden gezielte Aufgaben gestellt um das Gehör zu schulen. Die Studierende stellten mit erstaunen fest, dass es anfangs gar nicht so leicht war, mit etwas Übung die Geräusche jedoch lauter und deutlicher wurden.
Tasten
Der Tastsinn war als nächstes an der Reihe. Nachdem die einzelnen Teilnehmer einen Naturgegenstand in die Hand gelegt bekommen und diesen erfühlt hatten, gingen sie auf die Suche, ihren Partner mit dem gleichen Gegenstand zu finden. Hier stolperten die Studierende über die Schwierigkeit, die richtigen Worte für ihren Gegenstand zu finden. Aber auch hier wurden die Fähigkeiten schnell immer besser, sodass jeder seinen Partner fand, und sie zu Zweit in die nächste Aktivität konnten… „Bäume fühlen“. Zunächst sollte jeweils einer des Teams, blind in den Wald geführt werden und durch „Schwingungen“ erfühlen, wo der nächste Baum sich befand. In der zweiten Runde durften die Teilnehmer einen Baum mit ihren Händen erfühlen und diesen so kennenlernen. Die Studierenden berichteten begeistert davon, dass es zwar nicht immer leicht war, aber jeder Baum dennoch etwas Einzigartiges an sich hatte, an dem man ihn durchaus wiedererkennen würde.
Sehen
Die dritte Aktivität zielte auf das Sehen ab. Mit der Aktion „Umwelt im Umschlag“ wurden die Studierenden gezielt mit konkreten Aufgaben los geschickt um ihre Umwelt zu untersuchen und Gegenstände mitzubringen. Bei der Nachbesprechung und der Entwicklung weiterer möglicher und interessanter Fragen, brachte Noemi (ich hoffe an dieser Stelle, den richtigen Namen genannt zu haben) die Idee ins Spiel, den Geruchsinn mit anzusprechen. „Sammle mindestens 5 Dinge, die typisch nach Wald riechen.“ Gesagt, getan…
Gleichgewicht
Der letzte Sinn an diesem Tag war der Gleichgewichtssinn. Mit dem Spiel „Der schlafende Geizhals“ sollten die Studierende einem schlafenden Geizhals, welcher mit einer Wasserspritzflasche bewaffnet war, auf möglichst leisen Sohlen, die Schokolade, die vor ihm auf dem Waldboden lag, abnehmen. Gar nicht so einfach, wenn es bei jedem Schritt und Tritt knackst und raschelt.
Resumee
Nach einer schnellen Runde „Luftballon-Resümee“ war der Tag auch schon wieder vorbei.
„Ich habe heute gelernt, ohne zu merken, dass ich etwas lerne.“
29. Mai 2021 Obrteuringen, NSG Altweiherwiese
Treffpunkt an der Unterführung der Straße nach Bibruck unter der L 329 nach Meckenbeuren
1. Anfahrt von Ravensburg bis Oberteuringen auf der B33 bis Oberteuringen Ortsteil Hefigkofen
2. Dort am Gasthaus Adler nach links abbiegen Richtung Meckenbeuren
3. Der Straße folgen bis zur Abzweigung einer kleinen Straße links nach Bibruck
Quelle: Google Maps
Zum Naturschutzgebiet Altweiherwiese
Das 78 ha große Naturschutzgebiet Altweiherwiesen wurde 1981 vom Regierungspräsidium Tübingen ausgewiesen. Es liegt nordöstlich von Oberteuringen auf einer Meereshöhe von rund 450 m.
Im späten Mittelalter legten hier Mönche des Klosters St. Gallen durch Aufstau des Taldorfer Baches in Höhe der heutigen L329 einen Fischweiher an. Mitte des 18. Jahrhunderts wurde der Teich abgelassen und die feuchten Niederungen wurden bis Mitte des 20. Jahrhunderts als Streuwiesen genutzt. Heute werden die Wiesen von Naturschutz regelmäßig gemäht und das Mähgut wird entfernt. Dadurch wird das Mineralstoffangebot niedrig gehalten und das Aufkommen von Gehölzen verhindert.
Das umgebende Landschaftsschutzgebiet „Altweiherwiese und Taldorfer Bach“ soll das Naturschutzgebiet gegen störende Einflüsse von der Umgebung abschirmen.
Zur Landschaftsgeschichte
Auf einer topographischen Karte und noch deutlicher in einem digitalen Geländemodell erkennt man, dass das Naturschutzgebiet Altweiherwiese in der Fortsetzung eines Tals liegt, dass eine Verbindung zwischen Schussenbecken und der Niederung südlich des Gehrenbergs darstellt. Dabei handelt es sich um eine alte Schmelzwasserrinne, die am Ende der letzten Kaltzeit vor etwa 15.000 Jahren am damaligen Gletscherrand entstand. Dieses Rückzugsstadium des Gletschers, das sich einige Zeit hielt und kleine Endmoränen ablagerte, wird auch als Konstanzer Stadium bezeichnet. Die Schmelzwasserrinne war ein Teil der Ur-Argen, in der sich die Schmelzwasser vom nördlichen Gletscherrand sammelten und dem damals schon in Teilen existierenden Überlinger See zuflossen. In der Schmelzwasserrinne kam es immer wieder zu kleinen Aufstauungen, und Seenbildungen und darin zu Ablagerungen von Feinmaterial. Dadurch entstanden gegen das eiszeitliche Schottermaterial abgedichtete Bereiche, über denen es zu Torfbildung kommen konnte.
Ausdehnung des Rheingletschers vor ca. 15 000 Jahren. Die Schmelzwasserrinne am nördlchen Gletscherrand dieses als Kostanzer Stadium bezeichneten Gletscherrand-Verlaufs bildete die heutige Niederung des NSG Altweiherwiese (Zeichnung nach T. Gittner aus F. Beran 2002)
Heute wird die breite Talniederung von dem kleinen Taldorfer Bach durchflossen
Gräser
Gräser sind keine besonders auffälligen Pflanzen. Als Windbestäuber fehlen ihnen auffällige Blüten. Auf den ersten Blick kann man deshalb die verschiedenen Arten nur schwer unterscheiden. Bei genauem Hinsehen lassen sich jedoch meistens gute Bestimmungsmerkmale finden.
Zunächst probieren wir, ob man Grasarten auch blind unterscheiden kann, wenn man sie im Gesicht fühlt oder mit den Fingern ertastet. Es zeigt sich dass dies erstaunlich gut funktioniert, insbesondere, da bestimmte Merkmale, die man leicht übersieht – wie samtige Behaarung oder feine Grannen – sich ganz gut ertasten lassen.
Dann werden die verschiedenen Bestimmungsmerkmale der Süßgräser (Familie Poaceae) mithilfe eines Puzzles und realen Gräsern vorgestellt:
Blütenstand und Blüte
Als Einheit der Grasblütenstände gilt das Ährchen, das aus einer bis vielen Blüten bestehen kann. Diese Ährchen sind charakteristischerweise in einer Rispe angeordnet, seltener können die einzelnen Ährchen auch direkt an der Hauptachse sitzen, dann spricht man von einem Ährengras. Manchmal sind die Listen Äste sehr kurz, sodass der Blütenstand – obwohl er stärker verzweigt ist – wie eine Ähre aussieht (Ährenrispengras).
Jedes Ährchen hat an der Basis zwei Hüllspelzen, dann folgt vor jeder Blüte des Ährchens eine Deckspelze, eine Vorspelze und zwei Schwellkörper, die dafür sorgen, dass die Blütenteile bei der Reife auseinandergedrückt werden. Nach innen folgen drei Staubblätter und ein Stempel mit zwei fiederigen Griffelästen. Aus jeder Blüte schieben sich zunächst die Staubblätter mit langen beweglichen Fäden heraus und entlassen viele Pollen. Später entwickeln sich die federartigen Griffeläste mit den Narben die hervorragend zum auffangen der Pollen geeignet sind.
Halm
Die Sprossachse der Gräser ist meist unverzweigt und bildet mehrere auffällige Verdickungen. An diesen Knoten entspringen die wie eine Scheide den Stängel umfassenden Blätter. Nach einigen Zentimetern geht die Blattscheide in die Blattspreite über. An dieser Übergangsstelle finden sich entscheidende Bestimmungsmerkmale: das Blatthäutchen (Ligula) und die Öhrchen. Wenn man an einem Grashalm zieht, reißt er in der Regel an den Knoten, denn dort befindet sich wenig stabiles, teilungsfähiges Gewebe (interkalares Meristem). Im Unterschied zu den meisten anderen Pflanzen können sich die Sprosse wachsenden Gräser an den Knoten strecken und aufrichten, wenn sie vom Wind umgelegt wurden, oder sogar Wurzeln bilden. Das Bildungsgewebe an der Bruchstelle der Sprossachsen schmeckt süß.
Ausgewachsene Grashalme haben im Inneren oft einen Hohlraum (Trinkstrohhalm).
Verzweigungen
Die meisten Grasarten – eine Ausnahme bilden die Bambusse – verzweigen sich nur ganz nahe der Basis oder im Boden. Je nachdem, ob sich die Seitenzweige schnell nach oben krümmen oder ein Stück weit waagrecht wachsen, unterscheidet man Horstgräser, Rasengräser und Ausläufergräser.
Zum Schluss sammeln wir Gräser und ordnen sie entsprechend einem Merkschema.
Zwei weitere mit der Süßgräsern verwandte Familien sind die Sauergräser und die Binsengewächse
Zur Biberburg
Kurz vor der Brücke über den Taldorfer Bach nehmen wir den Weg rechts bergauf in den Wald. Der Weg verläuft etwas oberhalb des Baches. Wir beschäftigen uns mit einigen Pflanzen des Wegrandes zum Beispiel mit den Stickstoffzeigern (Zeigerpflanzen siehe unten) Klebriges Labkraut und Große Brennnessel. Zwischen den Brennnesseln wachsen Wald-Ziest und Hohlzahn, die ohne Blüten den Brennnesseln sehr ähnlich sehen. Andere Beispiele für Pflanzen-Mimikry (Nachahmen der Brennhaar-bewehrten Brennnesseln) sind Taubnesseln und Nesselblättrige Glockenblume.
Schließlich können wir durch das Unterholz die stattliche Biberburg erkennen. Ihre Eingänge liegen unter Wasser, aber die Wohnhöhle liegt über dem Wasser. Möglicherweise finden sich dort gerade junge Biber, denn normalerweise bringen Biber Ende April bis Anfang Mai ihre Jungen zur Welt. Sie bleiben bis zu einem Alter von 4-6 Wochen im Bau. Biber waren früher als Fastenspeise begehrt, da sie wegen ihres Schwanzes zu den Fischen gezählt wurden. Besonders wertvoll war der sehr dichte Biberpelz (23.000 Haare pro cm2, im Vergleich dazu kommt der Mensch nur auf ca. 200 Haaren pro cm2). Ein weiteres wertvolles, von Biber stammendes Handelsgut war das Bibergeil, ein Exkret, das in zwei etwa Hühnerei großen Blasen gesammelt und durch eine Ausführöffnung im Analbereich ausgeschieden wird. Dem Biber dient die Flüssigkeit der Fellpflege und der Reviermarkierung. Die Menschen nutzten sie wegen ihrer Inhaltsstoffe – zum Beispiel Hydroxybenzoesäuren und Abkömmlinge – als Medikament. Die Inhaltsstoffe oder ihre Vorläufer stammen vermutlich aus der pflanzlichen Nahrung des Bibers, zum Beispiel der Rinde von Weiden.
Der Biber ist das größte einheimische Nagetier. Mit seinem kräftigen Nagezähnen kann er große Bäume fällen. Ihre Zweige nutzt er einmal für den Bau seiner Burgen und Dämme, zum anderen dienen Knospen und Rinde als Nahrung. Für den Winter legt er Nahrungszweigdepots im Wasser an.
Zwischen Bast und Borke
Den nährstoffreichen Teil der Rinde, den Bast, der dem Assimilatetransport der Bäume dient, nutzen viele Tiere als Nahrung. Dazu zählen nicht nur Mäuse, Kaninchen, Hasen und Rehe, die an den Bäumen ihre Nagelspuren hinterlassen, sondern auch viel kleinere Tierchen, die Borkenkäfer. Sie gelten als Schwächeparasiten und die trockenen Sommer der vergangenen Jahre haben dazu geführt, dass insbesondere sehr viele Fichten von Borkenkäfern besiedelt wurden. Die rindenbrütenden Borkenkäfer legen ihre Eier unter der Rinde ab und die Larven fressen sich in langsam vergrößernden Gängen in den Bast. Diese Gänge kann man auf der Innenseite abgeschälter Rindenstücke sehr gut als Spuren erkennen. Form und Verlauf dieser Fraßgänge sind von Art zu Art unterschiedlich.
Gangsysteme des Buchdruckrs (Ips typographica) in Fichtenrinde, Teuringer Holz, 29.5.2021, Foto A. Winter
Außerdem gibt es sogenannte holzbrütende Borkenkäfer, deren Larvengänge in den Holzkörper hinein führen. Diese Käfer können sich von dem Holz nur mithilfe von Holz zersetzenden Pilzen ernähren, deren Hyphen die Gänge auskleiden und die sie fressen. Von außen kann man als Spuren dieser Käfer nur die Löcher im Holzkörper sehen. Größere Löcher stammen zum Beispiel von Bockkäfern. Wir finden die schon verlassene Puppenwiege eines Schrotbocks (Rhagium instructor), die aus kreisförmig ausgelegten Holzspänen besteht. Unter der abgestorbenen Rinde und unter den am Boden liegenden Rindenstücken kann man zahlreiche andere Tiere finden, die sich entweder von den organischen Abfallstoffen oder als Beutegreifer ernähren. Uns fallen vor allem zahlreiche Schließmundschnecken, einige Asseln, Tausendfüßler, Spinnen und kleinere sehr flinke Laufkäfer (wahrscheinlich Gattung Pterostichus) auf. Die meisten Borkenkäfer sind vermutlich ausgeflogen, aber wir finden noch einige Exemplare. Bei der Betrachtung durch die Becherlupen erkennen wir, dass an einigen Käfern Milben sitzen, welche die Käfer vermutlich nur als Transportmittel benutzen (phoretische Milben).
Weitere Beobachtungen
In dem Gebiet befinden sich zahlreiche alte Baumstümpe und abgestorbene Bäume. An mehreren Stellen solcher morscher (von Pilzen zersetzter) Stümpfe oder Stämme finden sich tiefe Hacklöcher, die vermutlich vom Schwarzspecht stammen, der regelmäßig im Gebiet zu hören und zu beobachten ist. Einen Pilzfruchtkörper identifizieren wir als Abgeflachten Lackporling (Ganoderma applanatum).
Hackloch eines Schwarzspechts (?) in einem morschen Eichenstamm (Weißfäule). Links sind die Reste eines Fruchtkörpers des Abgeflachten Lackporlings zu erkennen , Teuringer Holz 29.5.2021, Foto A. Winter
Ein typischer Pilzbefall an Rispengras-Halmen (rings um den Halm gehender weißer Belag) kann dem endophytische Pilz Epichloë typhina zugeordnet werden.
Nagelfluh-Kiesgrube
Der für mich mit dem Rollstuhl befahrbare Weg endet an einer verbreiterten Stelle mit steilen Wänden, die in der topographischen Karte als „Teuringer Holz“ markiert ist. Es handelt sich dabei um eine alte Kiesgrube, in der Interstadialer Nagelfluh abgebaut wurde, also verbackene Kiesablagerungen aus einer wärmeren Periode der Würm-Kaltzeit. In der Liste der Geotope im Bodenseekreis findet sich dazu folgende Beschreibung:
Aufgelassene Kiesgrube am Prallhang des Taldorfer Bachs, 1000 Meter östlich von Oberteuringen, an deren Südrand rund fünf Meter mächtige, zu Nagelfluh verfestigte interstadiale Schotter („alte Kiese von Oberteuringen“) zutage treten. Sie sind wahrscheinlich der „Laufenschwankung“, einem großen Rückzugsstadium zwischen Würm I und Würm II zuzuordnen. Der Aufschluss befindet sich am Südrand des Oberteuringer Eisrandtals, das erst später entstanden ist und hier mit einer Breite von etwa 500 Meter einen ehemaligen Gletscherrand markiert.
Ehemalige Kiesgrube im Teuringer Holz, 29.5.2021, Foto A. Winter
Riedwiesen
Botanisch besonders interessant ist die Riedfläche südlich der Straße, die nach Bibruck bzw. Wammeratswatt führt. Die Fläche wird von Naturschutz regelmäßig aber nicht jährlich gemäht. Wir können verschiedene Knabenkraut-bzw. Fingerwurz-Arten (Dactylorhiza majalis und. D. incarnata) finden, außerdem das Mittlere Zittergras (Briza media), Gelbe Segge (Carex flava) und Hirse-Segge (Carex panicea).
Breitblättriges Knabenkraut (Dactylorhiza majalis) und Mittleres Zittergras (Briza media), NSG Altweiherwiese, 9.5.2021, Fotos A. Winter
„Die Zeigerwerte sind Kurzbezeichnungen für das ökologische Verhalten, d. h. der Standortsbeziehungen der Pflanzen unter dem Einfluss zahlreicher Konkurrenten“ (Ellenberg 1991).
Pflanzen können Zeiger für bestimmte Standortfaktoren sein. Der Pflanzenökologe und Vegetationskundler Heinz Ellenberg, ordnete ab den 1950er Jahren den mitteleuropäischen Pflanzenarten aufgrund empirischer Erfahrungen Zeigerwerte für sieben verschiedene Umweltfaktoren zu, und zwar für die Standortfaktoren Licht (Lichtzahl), Temperatur (Temperaturzahl), Kontinentalität (Kontinentalitätszahl), Feuchtigkeit (Feuchtezahl), Stickstoff (Stickstoffzahl) und Bodenreaktion,pH-Wert (Reaktionszahl).
Als Beispiel seien hier die Zeigerwerte von drei Süßgräsern unterschiedlicher Standorte verglichen:
Art
Standort
L
T
K
F
N
R
Taube Trespe (Bromus sterlis)
Wegrand
7
7
4
4
X
5
Glatthafer (Arrhenaterum elatius)
Fettwiese
8
5
3
5
7
7
Mittleres Zittergras (Briza media)
Streuwiese bzw. Ried
8
X
3
X
X
2
Artenliste
(Zusammengestellt von Jenifer Friedrich und Anastaia Winter)
Gräser:
Süßgräser (Poaceae):
Glatthafer (Arrhenatherum elatius)
Kammgras (Cynosurus cristatus)
Wohlriechendes Ruchgras (Anthoxanthum odoratum)
Wolliges Honiggras (Holcus lanatus)
Gewöhnliches Rispengras (Poa trivialis)
Taube Trespe (Bromus sterilis) (Ruderalgras)
Weiche Trespe (Bromus hordaceus)
Wiesen-Fuchsschwwanz (Alopecurus pratensis)
Wiesen-Schwingel (Festuca pratensis)
Hunds-Quecke (Elymus caninus)
Fieder-Zwenke (Brachypodium pinnatum)
Mittleres Zittergras (Briza media)
Sauergräser (Cyperaceae):
Steife Segge (Carex elata)
Zittergras-Segge (Carex brizoides)
Wald-Segge (Carex sylvatica)
Gelbe Segge (Carex flava)
Hirsen-Segge (Carex panicea)
Binsengwächse (Juncaceae):
Feld-Hainbinse( auch Hain“simse“, aber „Simsen“ kommen auch bei der Familie Sauergräser vor) (Luzula campestris agg., Sammelart, unsere Pflanze war vermutlich L. multiflora))
Frau Margit Ackermann, Diplombiologin und Naturpädagogin und seit 2006 Mitarbeiterin des Naturschutzzentrums Wilhelmsdorf, gibt uns einen Einblick in Ziele und Aufgaben des Naturschutzzentrums und ihrer Arbeit. Das Zentrum wurde 1994 unter der privaten Trägerschaft des Schwäbischen Heimatbundes e. V. gegründet und mit Unterstützung des Landes Baden-Württemberg und der Gemeinde Wilhelmsdorf als Informationszentrum eingerichtet. 2016 übernahm die Stiftung Naturschutz Pfrunger-Burgweiler Ried die Trägerschaft. Neben den pädagogischen Aufgaben – Information der Öffentlichkeit Besucherangebote – geht es um Beobachtung und Dokumentation von Flora und Fauna, Durchführung von Artenschutzmaßnahmen, Organisation und Koordination von Pflegemaßnahmen einschließlich der extensiven Beweidung sowie der Flächenverwaltung und Verkehrssicherung. Derzeit läuft ein Verfahren zur Anerkennung des Pfrunger-Burgweiler Rieds als UNESCO-Biosphärenreservat.
Entstehungsgeschichte des Pfrunger-Burgweiler Rieds
In dem 2012 erstellten Ausstellungs- und Veranstaltungsgebäude versammeln wir uns vor einem Luftbild, in dem das Pfrunger-Burgweiler Ried mit Blick nach Süden – Bodensee und Alpen im Hintergrund – zu sehen ist. Vor 20.000 Jahren, beim Hochstand der letzten Kaltzeit (Würm-Kaltzeit) füllte eine Zunge des Rheingletschers das Tal zwischen den heutigen Orten Wilhelmsdorf und Ostrach. Beim Rückzug des Gletschers blieb am nördlichen Rand eine Endmoräne zurück (Äußere Würm-Endmoräne). In Höhe des heutigen Wilhelmsdorf kam es vor etwa 15.000 Jahren zu einem Stillstand der Gletscher-Rückentwicklung, eventuell auch einem weiteren Gletschervorstoß. In jedem Fall befand sich dort längere Zeit ein Gletscherrand und es lagerte sich Moränenmaterial ab (Innere Würm-Endmoräne). Das Schmelzwasser sammelte sich in dem Becken bis zur Endmoräne beim heutigen Ostrach, die den Abfluss nach Norden behinderte. Nach weiterem Abschmelzen wurde der Abfluss nach Süden durch die südliche Endmoräne blockiert, sodass ein Eisstausee erhalten blieb, in dem sich zunächst feines Tonmaterial ablagerte und den See nach unten abdichtete. Der Zufluss kalkreichen Wassers aus den umgebenden Höhenzügen, die teilweise aus interstadialen kalkreichem Nagelfluh aufgebaut sind, führte zur Ablagerung einer Seekreideschicht, das einsetzende Wachstum von Planktonalgen ließ eine erste Ablagerung mit hohem Anteilan organischen Material entstehen, die wegen ihrer braungrünen Farbe und der elastischen Eigenschaften als Lebermudde bezeichnet wird. Das mit der Klimaerwärmung zunehmende Pflanzenwachstum lieferte das Material für weitere Sedimente mit hohem organischem Anteil und Niedermoortorf. Im Laufe der Jahrtausende wurde der See dadurch immer flacher und aufgrund geringen Nährmineralgehalts und der hohen Regenmengen konnten sich auf den verlandenden Bereichen erste Torfmoose ansiedeln. Damit begann die Entwicklung zum Hochmoor.
Perspektivansicht des Pfrunger Zungenbeckens, des Rotach-Zungenbeckens und des westlichen Teils des Altshausener Beckens. Neben der Äußeren (blau gestrichelte Linie) und der Inneren Würm-Endmoräne (rote gestrichelte Linie) erkennt man auch eine Reihe von lokalen Moränenrücken (weiß gestrichelt). Das Pfrunger Zungenbecken entstand im Stadium der Äußeren Würm-Endmoräne. Weitere, jedoch wesentlich kleinere Zungenbecken des Stadiums der Äußeren Würm-Endmoräne entstanden bei Ilmensee (a) und im Altshausener Becken (b, c, d, e) Blickrichtung von Südosten nach Nordwesten (siehe Nordpfeil).
Der Illmensee und der Lengenweiher gehen auf Toteislöcher zurück, ursprünglich vom Gletscherrand abgetrennte Eispartien, die zunächst von Sediment überdeckt wurden und dann nach Abschmelzen wassergefüllte „Löcher“ bildeten.
Die Entwicklung bis zum heutigen Landschaftsbild wurde sehr stark durch menschliche Einflüsse geprägt. Nach der Gründung der Gemeinde Wilhelmsdorf durch württembergische Pietisten (Herrnhuter Brüdergemeinde) unter der Obhut des württembergischen Königs Wilhelm I. im Jahre 1824 wurde mit der Entwässerung und Kultivierung des Moorgebietes begonnen. Im 20. Jahrhundert wurde kurze Zeit ein intensiver industrieller Torfabbau betrieben. Diese Torfgruben sind bis heute als offene Wasserflächen erhalten.
Exkursion zum Hochmoorrest Eulenbruck
Unser Exkursionsweg -grün- (Karte aus einem Flyer des Naturschtzzentrums Wilhemsdorf)
Um einen Eindruck einer typischen Hochmoorlandschaft zu bekommen unternehmen wir eine Exkursion zum Eulenbruck. Wir folgen zunächst dem Asphaltssträßchen Richtung Lindenhof und biegen dann links ab und noch einmal links in den Kiefern-Moorwald mit einem dichten Unterwuchs aus Heidelbeeren (grüne Sprossachsen), an einigen Stellen auch Rauschbeeren (braune Sprossachsen). An einigen besonders feuchten Stellen kann man erste Ansiedlungen von Torfmoosen beobachten. Die Torfmoos-Pflänzchen haben einen typischen Aufbau aus Hauptachse (Stamm) und zu mehreren angeordneten dicht mit Blättchen besetzten Seitentrieben . Einige Seitenästchen laufen am Stämmchen herab und verleihen diesem eine dochtartige Struktur (Abb.).
Über einen Steg erreichen wir eine Aussichtsplattform, die den Blick auf einen relativ intakten Hochmoorbereich freigibt. Die geschlossene Torfmoosdecke, die wie ein Schwamm Wasser aufsaugt und festhält, wächst kontinuierlich nach oben, während die unteren Teile absterben und nur sehr langsam abgebaut werden. Dadurch entsteht kontinuierlich ein Zuwachs an Torf, in unserem Klima etwa 1 mm pro Jahr. Das bedeutet, dass in den 10.000 Jahren seit dem Ende der letzten Eiszeit maximal 10 m Torf gebildet werden konnte. Tatsächlich haben Bohrungen gezeigt, dass die Torfschichten unter dem Eulenbruck diese Mächtigkeit haben.
Durch einen Versuch demonstriert Margit Ackermann das schwammartige Wasseraufnahmevermögen von Torfmoosen. Aus einer Handvoll Torfmoose ausgepresstes Wasser wird in Sekundenschnelle wieder aufgenommen. Durch Auspressen gewinnt man allerdings nur einen Teil des in Torfmoosen gespeicherten Wassers, der größere Teil ist in einem Netz aus toten Wasserspeicherzellen in Blättchen und Stämmchen gespeichert, insgesamt das bis zu 30fache des Trockengewichts (Abb.). Da das gespeicherte Wasser nur aus dem Regen stammt, enthält es sehr wenig Mineralstoffe. Zudem haben Torfmoose die Fähigkeit, Mineralstoff-Kationen (K*; Ca++ u.a.) gegen H+-Ionen auszutauschen. Dadurch wird das Wasser stark abgesäuert. Wir messen in dem ausgepressten Wasser einen pH-Wert von ca. 4,0.
Torfmoose sind Regenwasserspeicher, sie ermöglichen die Hochmoorbildung. a) Torfmoospflänzchen, Habitus, b) Stämmchen quer mit Wasserspeicherzellen, Blättchen: c) Querschnitt und d) Aufsicht, e) räumliche Darstellung
Für Gefäßpflanzen sind Hochmoore deshalb ein sehr extremer Standort, weshalb hier nur wenige charakteristische Arten vorkommen. Die Heidekrautgewächse Moosbeere und Rosmarinheide können mithilfe ihrer Mykorrhizapilze und aufgrund ihres langsamen Wachstums hier gedeihen. Der Rundblättrige Sonnentau bessert seine Stickstoffversorgung durch Insektenfang und -verdauung auf (Carnivore Pflanze).
Blick von der Plattform Eulenbruck in den Hochmoorbereich mit Rundblättrigem Sonnentau (Drosera rotundifolia), Torfmoosen und Ranken der Moosbeere (Vaccinium oxycoccus) (Foto Rostan 25.7.2021)Aufbau eines Hochmoors in Mitteleuropa
Am Birkenaufwuchs und der stark aufkommenden Besenheide kann man erkennen, dass der Wassergehalt des Hochmoores einen kritischen Punkt erreicht hat. Die beiden letzten sehr trockenen Sommer konnten durch den diesjährigen bisher ziemlich regenreichen Sommer noch nicht ausgeglichen werden. Die Austrocknung bedeutet, dass die Zersetzung der organischen Materialien unter Sauerstoffeinfluss rascher voranschreitet . Das bedeutet nicht nur, dass das Moor nicht mehr in die Höhe wächst, sondern dass auch die tieferen Schichten abgebaut werden und der so über Jahrhunderte gespeicherte Kohlenstoff wieder als CO2 freigesetzt wird. Dies ist der Hauptgrund dafür, dass man angesichts der CO2-bedingten Klimaerwärmung Hochmoore unter besonderen Schutz gestellt hat.
Benennung verschiedener Feuchtbiotope
Insekten
Zurück am Naturschutzzentrum werden wir von schwärmenden Honigbienen empfangen. Wir entdecken eine Schwarmtaube an einem der Findlinge, die vor dem Naturschutzzentrum aufgestellt wurden. Der mit dem Naturschutzzentrum zusammenarbeitende Imker und Lehrbeauftragter der PH Weigarten, Herr Guggolz,wird informiert.
Nach wir eine kleinen Getränkepause und beschäftigen wir uns dann mit der Gruppe der Insekten, die aufgrund der zahlreichen Berichte über den dramatischen Rückgang ihrer Arten und Individuen im besonderen Interesse der Öffentlichkeit steht. Der starke Rückgang der Insekten-Biomasse ist mittlerweile so auffällig, dass er auch von Laien und der Entomologie fernstehenden Personen nicht mehr übersehen werden kann. Der starke Rückgang des Singvogelbestandes und insbesondere der Fledermäuse dürfte eine direkte Folge dieser Entwicklung sein. Auch die Ursachen sind nicht wirklich ein Geheimnis. Da ist erst einmal der hohe Pestizideinsatz in der Landwirtschaft zu nennen, andererseits aber auch der Verlust geeigneter Lebensräume mit einer vielseitigen Flora. Mit Blumenstreifen an Ackerrändern versucht man, einen Ausgleich zu schaffen. Da diese aus Saatmischungen stammenden Pflanzen häufig exotische, oft einjährige Arten enthalten stehen sie allerdings in der Kritik. Auch die Nähe zu den gespritzten Äckern und der Mangel an Brutmöglichkeiten und Nahrungspflanzen für die Larven der Insekten mindert den ökologischen Wert der bunten Ackerrandstreifen.
Eine wichtige Voraussetzung dafür, dass Insektenschutzmaßnahmen in der Öffentlichkeit breite Zustimmung finden, ist eine bessere Kenntnis der Insekten. Deshalb sind sie ein wichtiges Thema für den Biologieunterricht.
Dass man mit dem Thema durchaus Begeisterung wecken kann, beschreibt Dave Goulson in seinem Buch „Die seltensten Bienen der Welt“: „Es war ein sonniger Nachmittag gegen Schuljahresende im Juni 2009, und ich ging mit der Klasse meines ältesten Sohns Finn an der Newton Primary School Dunblane auf Insektenjagd….. Als wir am Wald waren, reichte ich den eifrigen Sieben- und Achtjährigen Netze und sonstiges Material und zeigte ihnen, wie man sie verwendet. … Einen Wiesenkescher zu öffnen, ist immer eine spannende Sache – wie bei den hübsch verpackten Geschenken unter dem Weihnachtsbaum weiß man nie, was Wunderbares drinsteckt. Unter lautem Ah und Oh sahen die Kinder zu, wie Scharen winziger Tiere – Ameisen, Spinnen, Wespen, Käfer, Fliegen und Raupen – aus dem Netz krabbelten, flogen und hüpften. Ich zeigte ihnen, wie man die kleinsten, empfindlichsten von ihnen in einen Exhaustor saugt. Dann verteilte ich eine Handvoll Becher, in denen jeder seinen Fang sammeln konnte, und die Kinder schwärmten aus, rannten durchs Unterholz, wedelten, kescherten und saugten nach Herzenslust, die Augen vor Aufregung weit aufgerissen. …“. Diese Beschreibung stammt aus dem Jahre 2009. Die Ergebnisse unserer Fangversuche waren deutlich schlechter – ein bisschen lag das vielleicht daran, dass die Witterung ziemlich schwül und wir durch unsere Exkursion schon etwas erschöpft waren, aber nicht nur. Ich habe mit Studierenden der PH Weingarten hier in Wilhelmsdorf zum ersten Mal 2017 Wieseninsekten gefangen und da waren unsere Ergebnisse deutlich vielseitiger. Auch die Fänge der parallel arbeitenden Wasserinsekten-Fanggruppe waren vergleichsweise dürftig (vgl. Artenliste).
Mittlerweile ist der Imker zu seinen Bienen gekommen und dadurch ergibt sich die Gelegenheit, aus erster Hand Informationen über dieses wichtigste Nutztier aus der Klasse der Insekten zu bekommen.
Die über 40.000 (immer noch?) mitteleuropäischen Insektenarten zu kennen, ist unmöglich, aber nicht so schwierig ist es, die wichtigsten 15-20 Ordnungen einheimischer Insekten zu erkennen und dann mithilfe eines Bestimmungsbuches auch einige Arten herauszufinden. Mir hilft dabei oft das mittlerweile leider schon längere Zeit vergriffene „Parays Buch der Insekten“ von Michael Chinery. Die über 2300 in diesem Buch mit sehr guten, treffenden Zeichnungen dargestellten Arten stellen nach meiner Erfahrung eine ausgezeichnete Auswahl dar, die zwar ursprünglich für Großbritannien getroffen wurde, aber auch für Mitteleuropa gilt.
Für Unterrichtszwecke hat Ulrich Kattmann eine Einteilung der Insekten in „Elfen, Ritter und Gaukler“ vorgeschlagen (vgl. Abb.)
Verwandtschaftsgruppen der Insekten
Die systematische Einteilung der Insekten nach Verwandtschaftsgruppen unterscheidet
ursprünglich flügellose Ordnungen, zu denen zum Beispiel Springschwänze und Silberfischen gehören,
Insekten mit unvollkommener Verwandlung, bei denen zwischen Larvenstadium und voll ausgebildeten Insekt (Imago) kein Puppenstadium eingeschoben ist, (zum Beispiel Libellen, Heuschrecken und Schaben) und
Insekten mit vollkommener Verwandlung (Ei – Larve – Puppe – Imago), zu denen zum Beispiel Käfer, Schmetterlinge und Hautflügler gehören.
Die meisten Bäume des Moorwaldes sind Wald-Kiefern (kenntlich an der braunrötlichen Färbung der oberen Stammbereiche), dazwischen stehen aber auch immer wieder Moor-Kiefern (mit durchgehend braunschwärzlichen Stämmen). Außerdem gedeihen hier Moor-Birken, die sich von Hänge-Birken durch die aufrechteren Zweige und die Behaarung der jungen Triebe unterscheiden.
Liste der beobachteten Pflanzenarten, chronologisch geordnet
(zusammengestellt von Darius Targan)
Name
Wissenschaftlicher Name
Echtes Mädesüß
Filipendula ulmaria
Echter Baldrian
Valeriana officinalis
Gewöhnlicher Blutweiderich
Lythrum salicaria
Sumpf-Hornklee
Lotus pedunculatus
Heidelbeere
Vaccinium myrtillus
Rauschbeere
Vaccinium uliginosum
Rosmarinheide
Andromeda polifolia
Hain-Gilbweiderich
Lysimachia nemorum
Wiesen-Wachtelweizen
Melampyrum pratense
Gewöhnliche Moosbeere
Vaccinium oxycoccos
Rundblättriger Sonnentau
Drosera rotundifolia
Roter Fingerhut
Digitalis purpurea
Kohl-Krazdistel
Cirsium oleraceum
Moor-Kiefer; Spirke, Moorspirke
Pinus mugo ssp. rotundata
Wald-Kiefer
Pinus sylvestris
Moor-Birke
Betula pubescens
Rasen-Schmiele
Deschampsia cespitosa
Wiesen-Lieschgras
Phleum pratense
Flutender Schwaden
Glyceria fluitans
Wald-Simse
Scirpus sylvaticus
Gewöhnlicher Dornfarn
Dryopteris carthusiana
Torfmoos
Sphagnum
Liste einiger beobachteter Tierarten
Säugetiere
Europäischer Biber (Castor fiber) -Biberausstiieg – Biberrutsche – am Weg begleitenden Bach
Vögel
Goldammer
Buchfink
Mönchsgrasmücke
Rabenkrähe
Insekten an Land
Brennnesselzünsler (Anania hortulata, Syn.: Eurrhypara hortulata): eingefaltete Brennnesselblätter lassen erkennen, dass sich hier Raupen eingesponnen hatten.
Landkärtchen (Araschnia levana) : Sommergeneration des Falters und Raupe
Die Anfahrt nach Wilhelmsdorf wird durch einige Gewitter und lokale Starkregen beeinträchtigt, wodurch einige Teilnehmer*innen abgeschreckt werden.
Der Abend beginnt im Unterrichtsraum des Naturschutzzentrums und er steht ganz unter dem Thema „Fledermäuse“. Diese außergewöhnliche Verwandtschaftsgruppe fliegender Säugetiere ist ein besonderer Schwerpunkt in der Arbeit des Naturschutzzentrums Wilhelmsdorf. Es geht dabei nicht nur um den Schutz und die Dokumentation der im NSG Pfunger-Burgweiler Ried vorkommenden Arten sondern auch um die Vermittlung von Wissen und den Abbau von falschen Informationen über die Flatterttiere. Frau Margit Ackermann führt uns in abwechslungsreicher und spannender Form in die Biologie der Fledermäuse ein. Dabei kommen verschiedene Spiele (Memory, Puzzle, Zuordnungsspiele), Anschauungsmaterial, ein Video und eine PowerPoint Präsentation zum Einsatz. Schließlich werden vier „Säuglinge“ der Zwergfledermaus (Pipistrellus pipistrellus)aus einer mit Tüchern abgedunkelten Box geholt und mithilfe einer Spezial-Minipipette und angerührter Milch für Hundewelpen gefüttert. Vor der Fütterung werden die Kleinen mithilfe einer Wärmflasche aufgewärmt um sie beweglicher zu machen.
Fütterung eines Zwergfledemaus-Babies (Foto S. Brendle)
Zum Abschluss – es ist mittlerweile 21:15 Uhr und beginnt langsam dunkel zu werden – wollen wir einige Fledermäuse in freier Natur erleben. Dabei helfen Fledermaus-Detektoren, durch welche die Ultraschall-Echolot-Signale der Fledermäuse in für uns hörbare Laute übertragen werden. In einem der beiden uns zur Verfügung stehenden Geräte werden nicht nur die Töne übersetzt, im Display wird auch noch ein Sonagramm (auch Sonogramm genannt) dargestellt, das den zeitlichen Verlauf der Lautäußerungen gegen die Frequenz aufzeichnet. Bei manchen Geräten wird auch noch die Lautstärke durch unterschiedliche Farben dargestellt.
Wir gehen vom Unterrichtsraum zum Bürogebäude des Naturschutzzentrums, an dessen Wänden verschiedene Fledermauskästen bzw. Fledermausbretter aufgehängt sind. Kaum stehen wir davor, wird schon die erste Fledermaus beobachtet, die Margit Ackermann mit dem Detektor als Zwergfledermaus identifiziert. Danach können wir längere Zeit einen großen Abendsegler am Himmel beobachten und mit dem Detektor hören. Wir wandern dann über die Straße und einen Weg zu einem größeren See und dort hören und sehen wir Zwergfledermaus, Wasserfledermaus und Großen Abendsegler, die meist dicht über die Wasseroberfläche fliegen.
Auf dem Rückweg taucht überraschend ein Biber vor uns auf und verschwindet später mit großem Platsch im Wasser.
Pflanzenfamilien erkennen
Je nach Zählung gibt es in Mitteleuropa mindestens 3000 verschiedene Pflanzenarten. Aber auch wenn viele davon selten sind oder nur in ganz bestimmten Gebieten vorkommen, so ist erscheint dem Laien auch die Anzahl von verschiedenen Pflanzenarten in der näheren Umgebung ziemlich unüberschaubar. Ganz allgemein gilt, dass man sich viele unterschiedlichen Objekte leichter merken kann, wenn man sie in Gruppen einteilt. Manche Pflanzenbestimmungsbücher nutzen hierfür zum Beispiel die Blütenfarben, andere Einteilungsmöglichkeiten sind der Lebensraum oder der Blühzeitraum. Eine Möglichkeit, die auch von Botanikern genutzt wird, ist die Einteilung nach der Verwandtschaft.
Die ersten Lebewesen auf der Erde sind vor mehr als 3 Milliarden Jahren entstanden. Alle heutigen Erdbewohner stammen von Ihnen ab und sind deshalb miteinander verwandt, aber nicht in gleichem Maße: Es gibt nähere und entferntere Verwandtschaften. Nahe verwandte und deshalb meist ähnliche Arten fasst man zu Gattungen, ähnliche Gattungen zu Familien zusammen, diese dann weiter zu Ordnungen, Klassen und Abteilungen.
Beispiel für die systematische Einordnung der Echten Nelkenwurz
Abteilung: Samenpflanzen
Klasse: Bedecktsamer Nacktsamer
Ordnung: Rosenartige Magnolienartige …
Familie:. Rosengewächse Brennnesselgewächse …
Gattung: Nelkenwurz Erdbeere …
Art: Echte Nelkenwurz Bach-Nelkenwurz …
Für das Bestimmen und Wiedererkennen von Pflanzen sind die Familien, in manchen Fällen auch die Gattungen, besonders wichtig und hilfreich. Wenn man die acht in den Kästchen vorgestellten, in der heimischen Flora häufigen Familien wiedererkennt, wird der Einstieg in die Artenkenntnis der Pflanzen leichter.
Quellen:
Beran, F. (2002): Die Entstehung des Natur- und Lebensraumes am nördlichen Bodenseeufer und um Oberteuringen. In: Gemeinde Oberteuringen (Hrsg., 2002): Teuringen. Ein Streifzug durch die Jahrhunderte, S.13-23.
Elenberg, H. (1991): Zeigerwerte der Gefäßpflanzen (ohne Rubus). In: Ellenberg, H. et al. (1991): Zeigerwerte von Pflanzen in Mitteleuropa. Scripta Geobotanica XVIII. Göttingen: E. Goltze
und die Materialien zu den früheren Exkursionen für die PH Weingarten ab 2016
Der Artikel erschien in dem mittlerweile vergriffenen UB-Heft 355 „Struktur und Funktion bei Pflanzen“ von 2010. Diese Fassung wurde an einigen Stellen erweitert, verändert bzw. aktualisiert.
Was sind Pflanzen?
Eine leichte Frage? – Aber was könnte man darauf etwa einem Außerirdischen antworten? Vielleicht: „Pflanzen sind Nichttiere.“ Also alle Lebewesen, die keine Tiere sind, sich nicht wie diese fortbewegen, fressen, mit ihren Sinnesorganen die Umwelt wahrnehmen und auf diese Umweltreize reagieren, Lebewesen die keine Nerven und Muskeln besitzen sind Pflanzen.
Wenn man das Inhaltsverzeichnis des „Strasburger, Lehrbuch der Botanik“ – seit mehr als hundert Jahren das klassische Lehrbuch der Pflanzenkunde – aufschlägt, entspricht der Inhalt dieser Vorstellung. In dem Lehrbuch werden Bakterien, Archäen, Pilze, Schleimpilze, Algen, Moose, Farne und Samenpflanzen behandelt – auch in der 37. Auflage von 2014. Einmal wird man bei genauerem Studium feststellen, dass die vorher aufgezählten „Vorurteile“ nicht alle aufrechterhalten werden können, zum zweiten sind die Unterschiede dieser Gruppen so groß, dass es nicht sehr sinnvoll erscheint, sie unter einem Oberbegriff zu fassen.
Vielleicht sollte man es deshalb mit einer positiven Definition versuchen: „Pflanzen sind grün“, sie enthalten Chlorophyll und sind damit zur Photosynthese in der Lage. Sie können aus Wasser und Kohlenstoffdioxid mithilfe von Lichtenergie und Mineralstoffen organische Stoffe wie Kohlehydrate, Lipide, Proteine und Nukleinsäuren aufbauen. Mit dieser Definition erreicht man, dass außer den Tieren auch die Pilze und die meisten Bakterien aus dem Pflanzenreich ausgeschlossen werden. Allerdings gibt es einige wirklich „echte Pflanzen“, die das Chlorophyll verloren haben und sich als Parasiten von anderen Pflanzen oder von Pilzen ihre Nährstoffe besorgen. Ihr Pflanzenstatus ist aber – wegen ihrer großen Ähnlichkeit zu verwandten grünen Pflanzen – kaum zu leugnen. Nicht sehr überzeugend an dieser Definition ist auch, dass damit sehr unterschiedliche Lebewesen, wie Blaugrüne Bakterien, viele chlorophyllhaltige Einzeller, mehrzellige, kugelige, fädige, flächige oder kompliziert strukturierte Algen sowie Moose, Farne und Samenpflanzen in einen Topf geworfen werden.
Deshalb erscheint es sinnvoller, als „Pflanzen“ Lebewesen zusammenzufassen, die nicht nur die Fähigkeit zur Photosynthese gemeinsam haben sondern auch noch weitere deutliche Ähnlichkeiten in Strukturen und Funktionen erkennen lassen. Solche Pflanzen haben einen dreidimensionalen Körper aus vielzelligen Geweben, der am Boden festgewachsen ist und sich in die Luft erhebt. Sie haben eine große Oberfläche, mit der sie im Stoffaustausch mit der Umgebung stehen und über die sie die für die Photosynthese nötige Lichtenergie auffangen können.
Entsprechend dem schon 1969 von Robert H. Whittaker vorgeschlagenen „Fünf-Reiche-System“ der Lebewesen werden wir uns in diesem Heft mit „Pflanzen im engeren Sinne“, also Moosen, Farnpflanzen und Samenpflanzen beschäftigen, dabei stehen die Bedecktsamigen Samenpflanzen im Mittelpunkt. Dies erscheint gerechtfertigt, da auch nach neuesten phylogenetischenn Vorstellungen ( ADL u. a. 2005) auf molekulargenetischer Basis die whittaker´schen Plantae eine monophyletische Gruppe sind.
Aus diesen Überlegungen ergibt sich eine funktionsbestimmte Definition für typische Pflanzen:
Pflanzen sind festsitzende, nachwachsende Lichtfänger mit Durchflusssystem
Festgewachsen in der Erde
Vom Lager zum Spross
Typisch für Pflanzen ist ein Vegetationskörper, der sich in die „Grundorgane“ Sprossachsen, Blätter und Wurzeln gliedert. Farnpflanzen und Samenpflanzen sind solche „Sprosspflanzen“ (Kormophyta). Die ursprünglichsten Pflanzen, die Moose, haben recht unterschiedliche Vegetationskörper, die jedoch in jedem Fall noch keine vollkommene Gliederung in Blätter, Sprossachsen und Wurzeln zeigen. Aber sie bilden wie Sprosspflanzen aus der befruchteten Eizelle einen von Schutzhüllen umgebenen Embryo, weshalb man beide zusammen auch als Embryophyta bezeichnet.
Abb. 1 Evolution der Pflanzen. Die für das Landleben besonders wichtigen neuen Merkmale der Sprosspflanzen sind am Beispiel von Rhynia dargestellt (Abb. teilweise aus Probst, W., Schuchardt,P. (Hrsg.): Biologie Lehrbuch für die S II, 584 S., Duden-Paetec, Berlin 2007)
Die ersten Pflanzen, die sich deutlich in den Luftraum erhoben, entstanden vor mehr als 400 Millionen Jahren, im Silur. Diese Urfarne bestanden, wie etwa in den Gattungen Cooksonia und Rhynia, aus gabelig verzweigten Achsen, die einige Dezimeter hoch werden konnten (Bell, Hemsley 2000, Abb. 2). Bei der Differenzierung in Sprossachsen, Blätter und Wurzeln blieben die ursprünglichen Gewebe – Bildungsgewebe, Abschlussgewebe, Grundgewebe, Leitgewebe, Festigungsgewebe – erhalten. Sie sind nur entsprechend den unterschiedlichen Funktionen der verschiedenen Grundorgane jeweils anders angeordnet (Kasten Grundorgane).
Grundorgane der Sprosspflanzen
Abb. 2 Grundorgane der Sprosspflanzen
Sprossachsen
Sprossachsen ähneln in ihrem Aufbau deutlich den ursprünglichen Achsen der Urfarne. Sie dienen dazu, die Assimilations- und Transpirationsorgane, die Blätter, im Luftraum zu positionieren. Dazu benötigen sie je nach Höhe eine größere oder kleinere Biegungsstabilität, die durch besondere Festigungsgewebe und Einlagerung von Lignin in die Zellwände erreicht wird. Außerdem müssen durch Sprossenachsen Stoffe über größere Distanzen transportiert werden (Wasser- und Assimilatetransport, Transport von Mineralstoffen, Hormonen usw.). Sie wachsen über Bildungsgewebe (Meristeme) an ihren Spitzen. Zunächst ruhende Meristeme in tiefer liegenden Bereichen (Knospen) führen, wenn sie aktiviert werden, zur Ausbildung von Seitensprossen (Zweigen).
Die Sprossachsen vieler Pflanzen sind zu sekundärem Dickenwachstum in der Lage, das von einem zylinderförmigen Bildungsgewebe, dem Kambium, ausgeht.
Blätter
Laubblätter sind Organe der Photosynthese und der Transpiration. Im Gegensatz zu Sprossachsen haben sie in aller Regel ein sehr begrenztes Spitzenwachstum, das später durch basales Wachstum ergänzt wird. Sie erreichen relativ schnell ihre endgültige Größe. Sie sind im typischen Fall in Stiel und Spreite gegliedert und zeigen eine deutliche Oben/Unten-Differenzierung der Gewebe als Angepasstheit an die Ausrichtung senkrecht zum Lichteinfall. Die Ausgestaltung der Blattorgane zeigt eine sehr große Vielfalt und viele Pflanzenarten kann man an der Form ihrer Blätter erkennen. Diese morphologische Vielfalt setzt sich in bestimmtem Umfang auch noch innerhalb einer Art und sogar innerhalb eines Individuums fort. Nicht selten kommt es zu einer großen Variationsbreite zwischen Jungendblättern und Altersblättern, Sonnenblättern und Schattenblättern usw. . ).
Typische Bildungen der Blattepidemis sind die Spaltöffnungen, die aus zwei meist bohnenförmigen Zellen bestehen, die eine Öffnung umschließen, die je nach äußeren und inneren Bedingungen geöffnet und geschlossen werden kann. Sie erlauben den Pflanzen eine Regulation des Gasaustausches, insbesondere der Transpiration von Wasserdampf und der Aufnahme von Kohlenstoffdioxid. Entsprechend ihrer Funktion als Transpirations- und Photosyntheseorgane sind Blätter in der Regel dorsiventral organisiert und senkrecht zum Lichteinfall hin ausgerichtet. Die Epidermis der Blattoberseite enthält keine Spaltöffnungen, nach innen schließt an die obere Epidermis ein Gewebe aus zylinderförmigen, dicht gepackten Zellen an. Dieses Palisadenparenchym enthält besonders viele Chloroplasten. Zur Blattunterseite hin folgt das lockere Schwammparenchym, zwischen dessen rundlichen Zellen große Interzellularen ausgebildet sind, die direkt mit den Poren der Spaltöffnungen in der unteren Epidermis in Verbindung stehen und so dem effektiven Gasaustausch dienen. Von diesem Grundaufbau eines Laubblattes gibt es allerdings sehr viele Abweichungen.
Wurzeln
Während Sprossachsen und Blätter in der Regel oberhalb der Bodenoberfläche gebildet werden, sind Wurzeln die im Boden liegenden Teile der Pflanzen. Neben der Stoffaufnahme dienen sie der Verankerung der Pflanzen und damit ihrer Standfestigkeit. Entsprechen dieser Funktionen tragen sie keine Blätter und bilden auch keine Spaltöffnungen aus. Die Anordnung des mechanischen Gewebes im Zentrum stärkt die Zugfestigkeit und nicht die Biegungsstabilität. Meist handelt es sich um sehr stark verzweigte Organe, die durch diese starke Verzweigung eine große Oberfläche bilden. Diese Oberfläche wird zusätzlich an den Wurzelspitzen durch sogenannte Wurzelhaare – Auswüchse von Wurzelhaut-(Rhizodermis -)Zellen – erhöht. Nur über diese vordersten Wurzelspitzen können die Pflanzen Wasser und Mineralstoffe aufnehmen. Bei ihrem Vordringen in den Boden werden die empfindlichen Wurzelspitzen von einer Wurzelhaube (Kalyptra) aus ständig nachwachenen Zellen geschützt.
Wie die Sprossachsen haben viele Wurzeln die Fähigkeit zum sekundären Dickenwachstum.
Sekundäres Dickenwachstum
Die kontinuierliche Verdickung der Achsenorgane ist für anhaltend wachsende große Pflanzen aus zwei Gründen wichtig:
zur Sicherung der Stabilität und
zur Gewährleistung der Transportkapazität
Im Gegensatz zu dem primären Dickenwachstum, das von den Apikalmeristemen ausgeht, ist für das sekundäre Dickenwachstum ein zylinderförmiges Meristem im Achsenorgan, ein Kambium, verantwortlich. Es kann entweder als Restmeristem des Vegetationspunktes erhalten bleiben oder sich sekunär aus bereits differenzierten Zellen neu bilden.
Bei Einkeimblättrigen Bedecktsamern ist sekundäres Dickenwachstum selten. Die sprichwörtlich schlanken Palmenstämme sind ein Beispiel dafür, dass selbst baumförmige Pflanzen ohne sekunäres Dickenwachstum auskommen können, allerings nur mit einem sehr ausgeprägten primären Dickenwachstum. Bei den Wurzeln umgehen viele Einkeimblättrigen das Problem dadurch, dass immer wieder neue sekundäre, sprossbürtige Wurzeln gebildet werden.
Abb. 3 Auf der linken Seite der schematisch dargestellte Aufbau einer typischen Sprosspflanze mit den Grundorganen Wurzeln, Sprossachsen und Blätter, auf der rechten Seite verschiedene Sonderformen: zwei chlorophyllfreie Parasiten, eine winzige Schwimmblattpflanze, eine Unterwasserpflanze fließender Gewässer mit starker Strömung (Blütentang – Podostemaceae) sowie eine Wüstenpflanze mit maximalem Verdunstungsschutz durch kugelige Oberfläche (Blütentang nach Tulasne 1852 aus Bell 1994)
Metamorphosen
Komplexe Evolutionsschritte sind nicht umkehrbar oder wiederholbar. Dies gilt jedoch nicht für den Funktionswandel. So kennt man im Pflanzenreich sehr viele Beispiele dafür, dass sich aus Sprossachsen (erneut) flache blattähnliche Organe gebildet haben und zwar in der Regel bei Pflanzen, die vorher aus anderen evolutionären Gründen ihre Blätter verloren haben. Am häufigsten kommt es zu einem solchen Blattverlust bei Pflanzen trockener Standorte (Xerophyten), die dadurch eine Verringerung der transpirierenden Oberfläche erreichen. Wenn später unter günstigeren Bedingungen der Besitz blattähnlicher Organe wieder von Vorteil wäre, kommt es jedoch nicht zu einer „Regeneration“ der verlorenen Blätter, vielmehr bilden sich aus Sprossachsenabschnitten „neue“ Blätter (Phyllokladien). Der Weg, von gefiederten Blättern wieder zu gnzrndigen Blattern zu kommen, führt über den Verlust der Blattspreite und die Verbreitrung des Blattstiels (Phyllodien, mehrfach bei der Gattung Acacia)
Besonders charakteristische Blattmetamorphosen, auf die schon Goethe aufmerksam gemacht hat, stellen die Blütenorgane dar. Unterirdische Sprossabschnitte können Wurzeln sehr ähnlich werden. Von echten Wurzeln unterscheiden sie sich jedoch oft durch kleine Schuppenblättchen und durch die Art der Verzweigung. Viele solcher unterirdischer Sprossachsen, die wie Wurzeln aussehen, sind uns recht vertraut, etwa die Rhizome (Erdsprosse) der Quecken, des Giersch oder der Winden.
Man spricht auch von „Metamorphosen“, wenn sich Grundorgane zu „neuen Organen“ umbilden, z. B. zu Dornen oder zu Ranken. Dabei sind Spross- und Blattdornen häufig, Wurzeldornen kommen nur selten vor. Dasselbe gilt für Sprossranken und Blattranken, Wurzelranken sind selten, Haftwurzeln (Efeu!) häufiger.
Abb. 4 Metamorphosen von Grundorganen
Emergenzen, Haare, Drüsen
Neben den Sonderstrukturen aus umgebildeten Grundorganen bilden Pflanzen auf ihren Oberflächen und auch im Inneren eine Vielfalt von Spezialstrukturen aus: Im Gegensatz zu Dornen sind Stacheln keine umgebildeten Grundorgane sondern Auswüchse oberflächennaher Gewebe. Haare entstehen aus einzelnen Oberflächenzellen, die sich jedoch mehrfch teilen können. Sie konnen rein mechanische Funktionen erfüllen, z. B. dem Verdunstungsschutz oder dem Lichtschutz dienen oder als kompakte Strukturen auch als Kletterhilfen. Besonders wirksame Fraßschutzeinrichtungen sind die Brennhaare der Brennnessel. Von manchen Haarzellen werden etherische Öle produziert (Drüsenhaare). Innere Drüsen sind oft mehrzellige Gebilde, die ihre Sekrete in interzellulare Hohlräume abscheiden. Sonderzellen in Geweben (Idioblasten) können der Stoffspeicherung oder der Ausscheidung dienen. Auch Harze und Milchsäfte können in speziellen Zellen im Inneren gebildet werden. Solche sogenannten sekundären Pflanzenstoffe sind für die Nutzung der Pflanzen als Heilkräuter oder Gewürze von Bedeutung.
Abb. 5 Emergenzen, Haare, Drüsen. 1 Papillenbildung bei Blütenblattepidermis vom Stiefmütterchen, 2 Klimmhaar des Hpfens, 3 Sternhaar einer Königskerze, 4 Brennhaar der Brennnessel mit mehrzelligem Sockel, 5 Drüsenhaar einer Minze,6 Drüsenhaar vom Lavendel, 7 Drüse vom Johanniskraut
Lichtfänger und Produzenten
Photoautotrophie
Das hervorragende gemeinsame Merkmal der Pflanzen ist, dass sie Lichtenergie zum Aufbau von organischen Bau- und Betriebsstoffen aus anorganischen Stoffen nutzen können (Photosynthese, vgl. z. B. UB 411, 328, 320, 249, 120, 35,). Sie fressen nicht, sie produzieren ihre Nährstoffe – Kohlenhydrate, Proteine, Lipide – selber und die Energie dazu liefert das Sonnenlicht. Dabei gelingt Pflanzen nicht nur die Assimilation von anorganischen Kohlenstoff sondern auch von Nitraten, Phosphaten und Sulfaten. Ähnlich wie Photovoltaikanlagen eine möglichst große dem Licht zugewandte Oberfläche benötigen, gilt dies auch für die „Photovoltaikanlage grüne Pflanze“, ihre Panele sind die grünen Blätter.
Diese primäre Produktion von organischen Stoffen macht sie – zusammen mit Algen und vielen Prokaryoten – zu Primärproduzenten, von denen die Stoffkreisläufe und Energieflüsse in allen Ökosystemen und in der gesamten Biosphäre ausgehen.
Dabei darf man allerdings nicht unterschlagen, dass die Pflanzen – genau wie alle anderen Lebewesen – für den lebenserhaltenden Energieumsatz Nährstoffe benötigen. Sie können diese Stoffe jedoch – im Gegensatz zu Tieren und vielen anderen Lebewesen – selbst herstellen, allerdings nur in Zellen mit Chloroplasten. Einige Pflanzen haben keine chloplastenhaltigen Zellen mehr und leben als Parasiten von anderen Pflanzen oder von Pilzen. Alle Pflanzen haben viele chloroplastenfreie Zellen und Gewebe, insbesondere ist das ganze Wurzelsystem normalerweise chloroplastenfrei und muss von den grünen oberirdischen Pflanzenteilen versorgt werden.
Primärstoffwechsel und Sekundärstoffwechsel
Wichtigstes Organell des aufbauenden (anabolen) Stoffwechsels sind die Chloroplasten, wichtigstes Organell des abbauen (katabolen) Stoffwechsels die Mitochondrien. Für die Lebensfunktionen grundsätzlich wichtige Stoffwechselwege fasst man als „Primärstoffwechsel“ zusammen. Von Zwischenprodukten (Metaboliten) des Primärstoffwechsels gehen gerade bei Pflanzen eine überaus große Anzahl von Art zu Art und von Verwandtschaftsgruppe zu Verwandtschaftsgruppe unterschiedlicher Stoffwechselwege aus, die man als Sekundärstoffwechsel bezeichnet . Bis heute kann man sich keine rechte Vorstellung von der Funktion der unübersehbaren Vielfalt dabei produzierter, meist flüchtiger organischer Substanzen machen. Ein Teil dieser Substanzen wirkt als Signalstoff, ein weiterer Teil dient der Abwehr von Fressfeinden. Zum Teil handelt es sich wohl nur um Ausscheidungen überflüssiger Stoffe. Global-ökologisch spielen solche gasförmigen Ausscheidungen von Pflanzen (VOCs von „volatile organic compounds“) eine wichtige Rolle, denn sie können sekundär organische Aerosole bilden, die die Wolkenbildung fördern (Probst 2009 in UB 349).
Stofftransport im Durchflusssystem
Voraussetzung für einen geordneten Stoffumsatz in einem Organismus ist, dass die Ausgangstoffe an den Reaktionsorten zur Verfügung stehen und dass die Endprodukte abtransportiert werden. Für die Stoffverteilung in den Pflanzen ist – anders als bei den durch ein Kreislaufsystem gekennzeichneten Tieren – der Wasserdurchfluss von den Wurzeln in die Blätter entscheidend. Auch Tiere müssen zwar ständig Wasser aufnehmen, doch im Vergleich zur im Körperkreislauf zirkulierenden Flüssigkeitsmenge ist diese Aufnahme gering – beim Menschen stehen 2-3 l täglicher Flüssigkeitsaufnahme 7000 bis 8000 l durch den Blutkreislauf bewegter Flüssigkeit gegenüber. Bei Pflanzen ist es genau umgekehrt. Auch hier gibt es in gewissem Umfang einen Kreislauf des Wassers zwischen Xylem und Phloem (Abb. 7), im Vergleich zum Wasserstrom, der vom Boden durch die Wasserleitungsbahnen bis zur Verdunstung in den Blättern führt, ist die dabei umgesetzte Menge aber sehr gering.
Abb. 6 Kreislaufsystem der Tiere und Durchflusssystem der Pflanzen
Die Notwendigkeit eines solchen Durchflusssystems hängt einmal mit der Aufnahme von Nährmineralien aus dem Boden zusammen. Für die Produktion organischer Nähr- und Baustoffe benötigen die Pflanzen außer Kohlenstoff, Wasserstoff und Sauerstoff vor allem die Elemente Stickstoff,Phosphor, Schwefel, Kalium, Calcium, Magnesium und Eisen, die als Mineralstoffionen mit der Bodenlösung aufgenommen und mit dem Wasserstrom an die Syntheseorte transportiert werden. Dies ist ein „positiver“ Grund für den Wasserdurchfluss.
Zum anderen ist die Kohlenstoffaufnahme nur über die oberirdischen Pflanzenteile aus der Luft möglich. Dabei ist das Kohlenstoffdioxid nur in sehr geringen Mengen in der Luft enthalten und um dieses wichtigste Gerüstelement aller organischen Verbindungen in ausreichender Menge aufnehmen zu können, ist eine große Oberfläche notwendig. Eine große aufnehmende Oberfläche bedeutet aber gleichzeitig eine große Oberfläche für die Wasserverdunstung. Dies ist ein „negativer“ (oder besser unvermeidlicher) Grund für den Wasserdurchfluss.
Eine große, dem Licht zugewandte Oberfläche ist – wie schon erwähnt – auch für die Photosynthese wichtig, zumal Pflanzen festgewachsen sind und dem Licht nicht hinterher laufen – höchstens ein bisschen hinterherwachsen – können. Die Verbindung mit der Wasserverdunstung ist dabei allerdings nicht ganz so eng wie bei der CO2-Aufnahme, da eine weitgehende Abdichtung der Oberflächen den Lichtfang nicht behindern würde.
Grundlage für alle Transportvorgänge in Pflanzen sind aktive oder passive Transporte durch Membranen, für Transporte über größere Entfernungen stehen spezielle Leitungsgewebe zur Verfügung.
Wasser- und Mineralstoffaufnahme in der Wurzel
Durch Diffusion bzw. Osmose wird Wasser an den Wurzelspitzen einmal in die Wurzelhaarzellen zum anderen kapillar in die Zellwände aufgenommen und osmotisch oder kapillar bis zu den Zellen der Endodermis geleitet. Dort verhindert eine Imprägnierung der seitlichen Endodermiszellwände einen weiteren kapillaren Wassertransport . Alles Wasser muss über die Membranen und das Plasma der Endodermiszellen geleitet werden. Dies gilt auch für die im Wasser gelösten Mineralstoffionen, die selektiv durch Zellmembranen aufgenommen werden, sowohl durch passiven als auch durch Stoffwechselenergie umsetzenden aktiven Transport.
Xylemtransport
Über die Endodermis gelangt das Wasser in die Leitbündel und in die bereits abgestorbenen röhrenförmigen Tracheen und Tracheiden. Treibende Kraft für den Weitertransport ist das negative Wasserpotenzial der Atmosphäre, das sich über die Spaltöffnungen, die Interzellularen der Blätter, die Kapillaren der Mesophyllzellwände und die Mesophyllzellen auf die Wassersäulen in den Leitungsbahnen auswirkt.
Dieser negative Druck kann jedoch nur zum Transport genutzt werden, wenn die Wasserfäden in den Tracheiden und Tracheen nicht abreißen. Dafür sind vor allem drei Sachverhalte entscheidend (Christian, Probst in UB 255,2010):
Die Kohäsionskraft zwischen den Wassermolekülen
Die Adhäsionskraft der Wassermoleküle an die Zellwände und Zellwandkapillaren
Die Oberflächenspannung in den Zellwandkapillaren am Ende der Wassersäule
Dies würde im Prinzip ausreichen, um Wasser weit höher als 100 m zu transportieren. Da es sich jedoch nicht um reines Wasser handelt, kommt es in den Leitungsbahnen über kurz oder lang trotzdem zur Gasblasenbildung (Cavitation). Nach Kanduč et al. 2020 hängt dies mit wasserunlöslichem Lipiden in den Wasserleitungsbahnen zusammen. Werden solche Cavitäten zu groß, reißt die Wassersäule ab. Dadurch wird die Stärke der theoretisch maximal tolerierbaren Unterdrücke von -1000 bar in reinem Wasser auf weniger als -100 bar reduziert..
Phloemtransport
Abb. 7 Druckstromtheorie – der kleiner Wasserkreislauf der Pflanzen
Auch für den Ferntransport der bei der Photosynthese produzierten Assimilate gibt es ein spezialisiertes Leitungssystem. Diese organischen Stoffe werden in wässriger Lösung über die Siebröhren bzw. Siebzellen des Phloems transportiert. Im Gegensatz zum Wasserstrom, der stets von den Wurzeln zu den Blättern fließt, kann die Assimilatelösung bedarfsabhängig in beiden Richtungen fließen. Über 90% des Siebröhrensaftes besteht aus Zuckern, v.a. aus Rohrzucker (Saccharose), außerdem sind Aminosäuren, Amide, Nucleotide (viel ATP) und organische Säuren enthalten. Motor des Transports ist der hohe osmotische Wert am Ausgangspunkt. Dadurch strömt Wasser in die Siebröhre ein. Der hydrostatische Druck führt dazu, dass Wasser am Ende der Leitung ausgepresst wird und Wasser von der Seite in die Siebröhren mit der höheren Konzentration nachfließt (Druckstrommodell). Für diesen Transport sind die siebartig durchbrochenen Querwände von Bedeutung.
Normalerweise werden die Assimilate in die wachsenden Meristeme an Spross- und Wurzelspitzen und in Speicherorgane (Knollen, Rüben) transportiert, überschüssiger Phloemsaft kann auch über besondere Drüsen (Nektarien) ausgeschieden werden. Vermutlich gehen die Nektardrüsen der Blüten auf solche Zucker-Ausscheidungsdrüsen zurück.
Blattläuse zapfen das Phloem an. Wenn sie ihre Rüssel aus den Siebröhren herausziehen, fließt der unter Druck stehende Phloemsaft noch einige Zeit nach und die überzieht die Blätter mit einem Zuckerfilm.
Durchlüftungssysteme
In den meisten pflanzlichen Gewebe schließen die Zellen nicht dicht aneinander, sie lassen vielmehr zwischen sich ein System aus Zwischenräumen (Interzellularen) frei, das letzten Endes über die oberflächlichen Spaltöffnungen mit der Außenluft in Verbindung steht. Der Stoffaustausch erfolgt über Diffusion. Dies gilt auch für Sumpf- und Wasserpflanzen und ihre unterirdischen bzw. untergetauchten Teile. Den langen Stängel der Seerosenblätter kann man als Schnorchel für das im Teichboden sitzende Rhizom auffassen.
Beim Mikrokospieren pflanzlicher Gewebe stört häufig der Luftgehalt in der Interzellularen, weil er im Nasspräparat wegen der anderen Lichtbrechung der Luft als schwarz umrandetes Objekt sichtbar wird. Deshalb ist es sinnvoll, größere Pflanzenorgane, z. B. Blätter oder Sprossachsen, bevor man sie mikroskopiert, in einer Saugflasche zu „entlüften“.
Nachwuchs
„Wachsen“ ist ein Begriff, der besonders mit Pflanzen verbunden wird, die man ja deshalb auch als „Gewächse“ bezeichnet. Wenn man Zweige abschneidet, wachsen sie nach, man kann Hecken hundertmal schneiden, Rasen tausendmal mähen und Feldhecken alle 8 Jahre „auf den Stock setzen“. Aus Knospen treiben diese gestutzten Pflanzen immer wieder neue Sprosse. Früher waren Niederwälder so ausdauernde Brenn- und Nutzholzlieferanten, nachwachsenden Rohstoffquellen. Eiben und Buchsbäume wurden von den Gärtnern der Barockgärten zu Skulpturen gestutzt.
Nachwuchs produzieren Pflanzen häufig asexuell , z. B. über Ausläufer, Ableger, Brutknospen, Brutzwiebeln. Nachwuchs kann aber auch sexuell hervorgebracht werden, bei ursprünglichen Pflanzen über einzellige Sporen, Samenpflanzen schützen ihren Nachwuchs im Samen, ähnlich wie Reptilien ihre Jungen in Eiern mit Eihüllen.
Die Entwicklung einer Pflanze lässt sich in 3 Phasen einteilen:
embryonale Phase
Wachstums und Reifephase
Reproduktive Phase und Seneszenz
Bei einjährigen Pflanzen kann dieser Lebenslauf innerhalb weniger Monate oder sogar Wochen ablaufen. Das wichtigste pflanzliche Versuchsobjekt, die Acker-Schmalwand (Arabidopsis thaliana) ist ein Beispiel dafür. Bei Bäumen wird die dritte Phase oft erst nach Jahren oder sogar Jahrzehnten erreicht und kann dann nahezu unbegrenzt anhalten.
Entwicklung des Embryos
Abb. 8 Entwicklung des Pflanzenembryos bis zur Keimung (nach Taiz, Zeiger, Physiologie der Pflanzen, Spektrum 2000)
Die ersten Entwicklungsschritte bis zur Anlage der Grundorgane laufen schon im Samen ab. Die Zygote streckt sich zunächst auf die etwa dreifache Länge, dann teilt sie sich inäqual. Aus der kleineren Zelle wird der Embryo (Keimling), die größere bildet den Suspensor, der den Embryo mit dem Nährgewebe der Samenanlage verbindet. Am Ende der Embryonalentwicklung steht das Herzstadium, in dem die Keimlingsstrukturen weitgehend festgelegt sind. An den beiden Endabschnitten der apikal-basalen Achse liegen die primären Meristeme, die als selbstregulierende Stammzellensysteme alle postembryonalen Strukturen der Pflanzen hervorbringen. Dazwischen liegen die Keimwurzel, das Hypokotyl (Sprossachse zwischen Wurzel und Keimblättern) und die Keimblätter. Die weitere Entwicklung führt im Prinzip immer wieder zu denselben Differenzierungen und damit auch zu denselben Strukturen (Wurzeln, Sprossachsen, Blätter). Diese Module können allerdings – z. B. bei der Blütenbildung (.s.u.) – erheblich variiert werden. Diese Entwicklung wird von Entwicklungsgenen gesteuert und von Umweltfaktoren und endogenen Faktoren moduliert.
Entwicklungskontrolle durch Genboxen
Alle pflanzlichen Entwicklungs- und Differenzierungsvorgänge werden von Phytohormonen gesteuert. Dazu müssen zunächst die Enzyme produziert werden, die für die Hormonsynthese notwendig sind. Gleichzeitig müssen auch Gene aktiviert werden, deren Produkte für die Empfindlichkeit einer Zelle gegenüber einem Hormon zuständig sind, z. B. die Rezeptormoleküle in der Zellmembran und die Proteine für eine mögliche Signalkette im Cytoplasma. Auch für die Steuerung der Translation im Zellkern und die anschließende Aktivierung oder Hemmung der Translationsprodukte können spezielle Genprodukte notwendig sein.
Alle Entwicklungsschritte werden dadurch möglich, dass von den Genen, die in allen Körperzellen vorhanden sind, einem genauen zeitlich–räumlichen Muster folgend immer nur ganz bestimmte Gene exprimiert werden. Dieses Muster der Entwicklung wird durch eine Hierarchie von Kontrollgenen möglich, die jeweils für Transkriptionsfaktoren codieren, die andere Gene an- oder abschalten können. Für die DNA-Bindung dieser als Transkriptionsfaktoren wirkenden Genprodukte ist eine besondere Proteindömäne verantwortlich, der einem bestimmten Genabschnitt entspricht, der als „Box“ bezeichnet wird. Bei diesen Boxen handelt es sich um sehr konservative Genabschnitte, die sich über lange Evolutionsabschnitte nicht verändert haben und die zum Teil allen Eukaryoten gemeinsam sind (Seyffert 2003, S. 699 ff)
Organidentitätsgene steuern die Bildung der Blütenorgane
Eine für Pflanzen besonders bedeutende Proteindomäne ist die MADS-Box. Mittlerweile kennt man über 100 verschiedene MADS-Box-haltige Transkriptionsfaktoren, die in allen Reichen der Lebewesen vorkommen. Der Name stellt die Anfangsbuchstaben von vier Genprodukten dar, von denen zwei bei Pflanzen, eines bei der Bäckerhefe und eines beim Menschen gefunden wurden. Erste erfolgreiche Untersuchungen zur Wirkung von Kontrollgenen bei Pflanzen wurden an MADS-Box-Genen durchgeführt, die für die Entwicklung des Blütenbereiches wichtig sind und Organidentitätsgene genannt wurden.
Als Blütenorgane bezeichnet man die wirtelig angeordneten Teile einer Blüte, die von außen (bzw. unten) nach innen (bzw. oben) als Kelchblätter, Kronblätter, Staubblätter und Fruchtblätter (meist verwachsen zum Stempel) bezeichnet werden. Schon Goethe war aufgefallen, dass es bei der Blütenbildung ab und zu „Verwechslungen“ zwischen den Blütenwirteln kommen kann, dass z. B. aus Staubblättern Kronblätter werden und „gefüllte“ Blüten entstehen.
Die Entwicklung zu einer Blüte beginnt am Vegetationspunkt einer Sprossachse. Das Meristem an der Sprossspitze bildet – induziert durch äußere oder innere Faktoren – keine Laubblattanlagen mehr, sondern Anlagen von Blütenorganen. Dabei kann man drei Hierarchieebenen von Genen unterscheiden. Auf der untersten Ebene geht es um die Ausbildung der verschiedenen Blütenwirtel und dabei werden die Organidentitätsgene wirksam (Abb. 9):
Gen A wird in den beiden äußeren Wirteln exprimiert, die zu Kelch- und Kronblättern werden.
Gen B wird in den Wirteln zwei und drei exprimiert, die Kronblätter und Staubblätter bilden.
Gen C wird in den beiden inneren Wirteln exprimiert, aus denen Staubblätter und Fruchtblätter hervorgehen.
Die Wirkungsweise der Organidentitätsgene kann man sich so vorstellen, dass die Gene A, B und C jeweils nur für eine Untereinheit eines aus zwei Untereinheiten zusammengesetzten Transkriptionsfaktors codieren: A-A, A-B, B-C, C-C. Man kann in diesem Fall von einer kombinatorischen Genregulation sprechen. Die Zusammensetzung des Dimers entscheidet darüber, welche anderen Gene von dem Transkriptionsfaktor aktiviert werden. Besteht im Beispiel ein Dimer nur aus den beiden Untereinheiten A, werden Kelchblätter produziert, besteht es aus A und B, bilden sich Kronblätter aus, besteht es aus B und C, werden Staubblätter gebildet und besteht es aus zwei C´s, entstehen Fruchtblätter.
Wenn nun durch eine Mutation der Promotor, der normalerweise Gen C aktiviert, an Gen A gekoppelt wird, bilden sich keine Staub- und Fruchtblätter, sondern nur Kelch- und Kronblätter aus. Dieses Grundmuster wird durch weitere Organidentitätsgene noch etwas differenziert, außerdem gilt es nur für einen Teil der Bedecktsamer.
Abb. 9 Blühinduktionsgene steuern die Blütenbildung
Licht spielt als entwicklungssteuernder Faktor bei Pflanzen eine entscheidende Rolle. Lichtabhängige Entwicklungsvorgänge (Photomorphogenese) können von Blaulicht oder Rotlicht abhängen.
Keine Vorurteile gegenüber Pflanzen !
Gängige Vorurteile sprechen Pflanzen tierliche Fähigkeiten ab: Pflanzen sind bewegungslos, sie haben keine Sinnesorgane, kein Skelett und keine Muskeln, sie haben kein Nervensystem und sie können nicht Kommunizieren und Kommunikationen verarbeiten („Denken“). Gerade bei Kindern führen solche Vorstellungen dazu, dass Pflanzen nicht unbedingt als Lebewesen eingestuft werden, deshalb sollen sie etwas ausführlicher widerlegt werden.
Bewegungslos?
Wenn man die reife Frucht eines Springkrauts anfasst, spürt man eine Reaktion zwischen den Fingern, die fast an einen Muskel erinnert. Die Frucht platzt auf, indem sich die Fruchtklappen nach innen rollen und die Samen werden ausgeschleudert. Allerdings liegen dieser Bewegung keine Muskelzellen und auch kein Nervengewebe zugrunde, denn diese spezialisierten Zellen kommen bei Pflanzen nicht vor. Trotzdem sind Reaktionen auf Umweltreize und die Verarbeitung solcher Reize möglich und in einigen Fällen führt dies auch zu auffälligen und schnellen Bewegungen.
Krümmen, Klettern, Öffnen und Schließen – Wachstumsbewegungen
Viele beobachtbare Bewegungen von Pflanzen, die auch Reaktionen auf Umweltreize darstellen können, gehen auf Wachstumserscheinungen zurück. Wenn sich in der zylinderförmigen Sprossachse einer Pflanze die wachsend Zellen auf der einen Seite stärker strecken als auf der anderen Seite, kommt es zu einer Krümmung. Wenn diese Streckungen periodisch um die Achse herum stattfinden, kommt es zu Windebewegungen. So kann das Sprossende einer Zaunwinde oder einer Bohnenpflanze kreisende Bewegungen ausführen, die wie das Suchen nach einer Unterlage aussehen. Ist diese Unterlage gefunden, wird sie relativ schnell umwunden, denn die Pflanze kann den Widerstand fühlen. Noch komplizierten wird es bei der Aufrollbewegung von Ranken. Auch hier wandert das Streckungswachstum um die Sprossachse, aber – da die Ranken am Vorder- und am Hinterende fixiert sind – muss mindestens ein Umkehrpunkt eingebaut werden. Bei der Steuerung dieser Bewegungsvorgänge von Windepflanzen spielen sowohl endogene Rhythmen als auch Umweltreize, z. B. Berührungsreize, eine wichtige Rolle.
Noch weiter verbreitet ist die Hinwendung pflanzlicher Sprossachsen zum Licht, also eine einfache Krümmungsbewegung in Richtung einer Lichtquelle. Umgekehrt zeigen Wurzeln oft eine Krümmung vom Licht weg. Solche Wachstumsbewegungen, die von einem Außenreiz ausgelöst werden, nennt man auch Tropismen und je nach Reiz spricht man von Photo-, Geo- oder Hydrotropismen, die positiv oder negativ sein können oder auch das Mittel zwischen zwei Reizeinflüssen einhalten können (Plagiotropismus), z. B. beim horizontalen Wachstum von Seitenzweigen. Nicht selten führen Pflanzenorgane richtiggehende Wachstumsprogramme durch. Die Blütenstiele von Mohnpflanzen z.B. krümmen sich vor dem Aufblühen stark ein, bei Auflühen strecken sie sich wieder. Eine ähnliche Abfolge von Krümmung und Streckung kann man bei den Blütenstände zu flach nichts beobachten.sdd
Vergleicht man solche Wachstumsbewegungen von Pflanzen mit Bewegungen von Tieren, so ergeben sich deutliche Unterschiede. Insbesondere sind diese auf Wachstumsvorgänge beruhende Bewegungen alle sehr langsam, sie können von uns nur indirekt wahrgenommen werden und sie sind eher mit den Form- und Proportionsveränderungen vergleichbar, die im Laufe der Keimesentwicklung von Tieren auftreten.
Die zweite große Gruppe pflanzlicher Bewegungen beruht auf Änderungen des Zellbinnendrucks von Pflanzenzellen, des sogenannten Zellturgors, der in der Regel durch den unterschiedlichen Wassergehalt der Zentralvakuolen der Pflanzenzellen zustande kommt. Dieser wird wiederum über den osmotischen Wert gesteuert.
Für fast alle Pflanzengruppen sind Spaltöffnungen charakteristisch. Das sind von zwei Schließzellen umgebene Poren in der Außenhaut (Epidermis), die in Abhängigkeit von äußeren und inneren Reizen geöffnet und geschlossen werden können. Ist der Turgor der bohnenförmigen Schließzellen hoch, bilden sie eine nahezu kreisförmige Gestalt, die in der Mitte einen Porus offen lässt. Ist der Turgor niedriger, sorgt der Druck des übrigen Gewebes dafür, dass die Schließzellen aneinandergepresst werden. Dies ist die klassische Bewegungsreaktion, es gibt viele Abwandlungen und Sonderformen, die durch die besondere Gestalt der Schließzellen und ihre Anordnung in der Epidermis zustande kommen. Turgorschwankungen sind aber in jedem Fall der Motor der Bewegung. Ursache für diese Turgorschwankungen sind Schwankungen im Ionengehalt, insbesondere im Kaliumionengehalt der Schließzellvakuolen und dieser Ionengehalt kann durch aktive Pumpmechanismen verändert werden. Die Regulation der Spaltöffnungsbewegung sorgt dafür, dass die Pflanzen mit der schwierigen Aufgabe zurecht kommen, in der trockenen, kohlenstoffdioxidarmen Atmosphäre weder zu verhungern noch zu verdursten.
Spektakulärer, da ohne optische Hilfsmittel sichtbar, sind andere Turgorbewegungen von Pflanzen. Da wären so auffällige Beispiele wie die „Schamhafte Mimose“ oder Sinnpflanze (Mimosa pudica) oder die Venusfliegenfalle (Dionaea muscipula) zu nennen. Die Reizreaktionen dieser Pflanzen erinnern wirklich an nerven- und muskelgesteuerte Bewegungen von Tieren. Für die Bewegungen sind nicht nur einzelne Zellen, wie bei den Spaltöffnungen, sondern ganze Gewebe verantwortlich, deren Turgor aktiv verändert werden kann. Dabei kommt die schnelle Bewegung in der Regel durch einen plötzlichen Turgorabfall zustande, die anschließende „Regeneration“, die mit einem Aufbau des alten Turgorzustanden verbunden ist, dauert etwas länger.
Ähnlich wie das Öffnen und Schließen einer Tür werden Turgorbewegungen zwar oft von Außenreizen bewirkt, sie laufen aber unabhängig von der Richtung der Reizquelle ab. Solche Bewegungen werden als Nastien bezeichnet
Neben reversiblen Turgorbewegungen kennt man irreversible. Hierzu zählt das oben angesprochene Beispiel des Springkrauts. In den schmalen zylinderförmigen Zellen der äußeren Fruchtwand wird ein hoher Turgordruck aufgebaut, der schließlich zu einem Aufreißen der Frucht an präformierten Nähten zwischen den Fruchtblättern führt. Sind die Fruchtblätter erst voneinander getrennt, können sich die zylinderförmigen Zellen ausdehnen und dadurch werden die Fruchtblätter nach Innen eingerollt. Dabei spielt allerdings auch noch die elastische Zellwand eine Rolle. Wenn sich die Zellen der Außenwand erst einmal abgerundet haben, ist eine Rückkrümmung er Fruchtklappen nicht mehr.
Verbiegen und Verdrehen – Quellungsbewegungen
Für das Funktionieren von Pflanzen sind auch Bewegungen von Bedeutung die keine physiologischen sondern rein physikalische Ursachen haben, hierzu zählen Quellungs- und Kohäsionsbewegungen. Entscheidend für diese Bewegungen sind Zellulosestrukturen (Abb. 3). Zellulose ist aus fädigen Molekülen aufgebaut, die zu sogenannten Mizellen zusammengefasst sind. Diese Zellulosesemizellen können in unterschiedlicher Richtung in eine Zellwand eingelagert sein, ähnlich wie die Faserstrukturen im Papier. Zwischen die Mizellen kann Wasser eingelagert werden. Dies führt zu einer Ausdehnung und zwar stärker quer zur Faserrichtung als in Faserrichtung. Sind die Faserrichtungen in zellulosehaltigen Pflanzenstrukturen geordnet aber in verschieenen Schichten unterschiedlich, so kann dies beim Quellen zu Krümmungsbewegungen führen. Bei den Schuppen von Nadelholzzapfen zum Beispiel, aber auch bei vielen Fruchtklappen oder -zähnchen sind die Zellulosemizellen außen vorwiegend quer und innen längs angeordnet. Die stärkere Außenquellung bei Wassereinlagerung führt dazu, dass sich Nadelholzzapfen bei Feuchtigkeit schließen und bei Trockenheit durch nach außen Krümmen der Schuppen wieder öffnen. Dadurch wird erreicht, dass die Samen nur bei Trockenheit ausfallen können, was für deren Windverbreitung von Vorteil ist. Weitere auffällige Beispiele für solche Quellungsbewegungen zeigen viele Öffnungsklappen von Früchten oder die Peristomzähnchen an den Kapseln von Moossporogonen.
Je nach Anordnung der Micellen kann bei diesen Bewegungen nicht nur zu Krümmungen sondern auch zu Schraubungen kommen. Besonders einrucksvoll ist die lange, wie ein Drillbohrer wirkene Fruchtklappe des Reiherschnabels.
Auch solche Quellungsbewegungen können mit dem Bruch von präformierten Nähten verbunden sein. Bei vielen Schmetterlingsblütlern öffnen sich die Hülsen nahezu explosionsartig, beim Blauregen (Wisteria sinensis) zum Beispiel mit einem richtigen Knall. Dies kommt dadurch zustande, dass bei Austrocknung sich allmählich eine Spannung aufbaut, die schließlich zum Aufreißen an präformierten Nahtstellen führt. Oft kann man dieses Aufreißen als kleinen Knall wahrnehmen (z. B. bei Lupinen, beim Blauregen oder bei der Blatterbsen-Wolfsmilch).
Bei dem Aufreißen von Farnsporangien, spielt die Köhäsion der Wassermoleküle eine wichtige Rolle.
Von einem Ort zum andern
Pflanzen können nicht aktiv kriechen, laufen, schwimmen oder fliegen, sich zum Beispiel gerichtet auf eine Reizquelle (Taxie) zubewegen oder einer Spur folgen. Passive Ortsbewegungen sind aber möglich. So sind viele Wasserpflanzen nicht festgewachsen. Sie fluten frei an der Oberfläche oder untergetaucht im Wasserkörper und werden von Strömungen mitgetragen. Nur so konnte sich die Wasserhyazinthe (Eichhornia crassipes) – ursprünglich im Amazonasgebiet zuhause – über fast alle tropischen und subtropischen Gewässer verbreiten und die Kanadische Wasserpest (Elodea canadensis) hätte andres nicht in kurzer Zeit zu einer die Schiffahrt bedrohenden „Pest“ in europäischen Gewässern werden können.
Einjährige Wüsten-oder Steppenpflanzen können, wenn sie nach der Samenreife absterben, zu „Wüstenrollern“ werden, die vom Wind beträchtliche Strecken über den Steppenboden gerollt weren und dabei die Samen allmählich ausstreuen.
Dies schafft den Übergang zu den Pflanzenteilen, die auf Fortbewegung spezialisiert sind, weil sie der Ausbreitung dienen sollen. Sie werden Diasporen genannt. Diasporen können Sporen, Samen, Früchte, Teilfrüchte und vegetative Ausbreitungseinheiten wie Brutknospen, Brutzwiebeln, Spross- und Wurzelknollen oder Butkörper sein. Triebkräfte bzw.Transportmittel für die Ortsbewegung sind Wind, Wasserströmung, die Schwerkraft, Tiere und der Mensch. Dabei wird in der botanischen Literatur streng zwichen Ausbreitung, dem Fortbewegungsvorgang einer Diaspore, und der oft daraus resultierenden geografischen Verbreitung einer Pflanzenart unterschieden.
Knochenlos?
Die Bewegungen der Tiere kommten i. A. durch das Zusammenspiel von Muskeln und Skelett zustande. Dabei kann es sich um ein inneres Knochenskelett (Wirbeltiere), einen äußeren Skelettpanzer (Indsekten und andere Panzerhäuter – Ecdysozoa) oder auch ein Hydroskelett (Regenwurm und andere Würmer) handeln. Pflanzen haben sowohl ein den Knochen entsprechendes Innenskelett, das aus Geweben mit verdickten Zellwänden besteht, als auch ein Hydroskelett.
Tierzellen haben keine Zellwände, die harten Skelettelemente bilden sich aus Abscheidungen der Zellen in den extrazellulären Raum. Demgegenüber besitzt jede Pflanzenzelle eine feste Zellwand, die vor allem aus langkettigen Polysacchariden, insbesondere aus Zellulose, besteht. Es können aber weitere Stoffe eingelagert oder aufgelagert sein, z. B. Lignin bei verholzten Zellwänden oder Siliziumdioxid bei Gräsern und Schachtelhalmen. Pflanzenzellen machen in der Regel nach ihrer Bildung eine lange Phase der Volumenzunahme durch, wobei eine starre Zellwand sehr hinderlich wäre. Deshalb sind die Polysaccharidfasern in den Zellwänden junger Pflanzenzellen noch wenig fixiert und der Volumenzunahme folgend können immer neue Fasern eingebaut werden. Erst wenn die Zelle ihre endgültige Größe erreicht hat, kommt es zu einer zunehmenden Stabilisierung und Verfestigung der Wände, die vor allem durch einen regelmäßigen Wechsel in der Textur der Zellulosefibrillen und zunehmenen Querverbindungen zwischen den Fibrillen erreicht wird.
Durch die Bildung von Zellen mit sehr dicken Zellwänden können mechanisch stark belastbare Gewebe entstehen. Dabei kann es sich um lange Zellfasern handeln, die vor allem Zug- und Dehnungsstabilität bewirken und die von der Textilindustrie auch als „Pflanzenfasern“ genutzt werden (Baumwolle, Hanf, Lein, Jute, Sisal usw.) oder um isodiametrische Steinzellen, die vor allem Druckbelastungen standhalten. Durch die kompakte Anordnung von Zellen mit stark verdickten und verholzten Zellwänden kann eine hohe Druck- und Biegungsstabilität erreicht werden.
Besonders eindrucksvoll sind die kompakten Holzkörper der großen Bäume, die allerdings zum großen Teil aus abgestorbenen Zellen bestehen. Der lebende Baum bildet nur einen hautartigen Überzug über dem toten Holzskelett. Es gibt aber auch beeindruckene Beispiele für Leichtbauweisen. Viele Pflanzenkonstruktionen zeigen, wie mit möglichst geringem Materialaufwand möglichst große Stabilität oder Festigkeit erzeugt werden kann, z. B. reißfeste Blätter, biegungsstabile Grashalme, oder tragfähige Säulenkakteen.
Typisch für Pflanzenzellen sind außerdem Vakuolen mit wässerigem Inhalt, die bei ausgewachsenen Zellen meist den größten Anteil des Zellinneren ausmachen (Zentralvakuole). Je höher der Gehalt der Vakuole an osmotisch wirksamen Substanzen, je niedriger also ihr Wasserpotenzial, desto mehr Wasser wird von außen angezogen und desto größer wird der Binnendruck des Zellkörpers gegen die Zellwand, den man auch als Turgor bezeichnet. Für die Stabilität eines Pflanzensprosses ist dieses durch den Turgor gebildete Hyroskelett von großer Bedeutung. Dies kann man daran erkennen, dass Pflanzen bei Wasserverlust „verwelken“. Sie sind dann nicht mehr in der Lage, ihr eigenes Gewicht zu tragen: Ihre Blätter hängen schlaff herab und sie biegen sich zum Boden.
Vakuolen dienen aber auch als Stoffreservoir für die Pflanzenzellen. Hier können organische Säuren gespeichert werden, ebenso wasserlösliche Farbstoffe. Es kann auch zum Ausfallen schwer löslicher Verbindungen in Vakuolen kommen, die dann mit dem Absterben der entsprechenden Pflanzenteile, z. B. der Blätter, auf diese Weise ausgeschieden werden. Eine wichtige Funktion der Vakuolen ist auch die der Wasserspeicherung. Besonders große Vakuolen kommen in den Früchten vor und wir genießen den leckeren Vakuoleninhalt, wenn wird die saftigen Früchte essen oder die ausgepressten Fruchtsäfte trinken.
Sinnenlos?
Pflanzen haben keine richtigen Sinnesorgane wie Augen und Ohren, sie sind aber durchaus in der Lager, physikalische und chemische Umweltreize wahrzunehmen, insbesonere Licht und Schatten, Schwerkraft, Erschütterungen und Berührungen, Wärme (bzw. Temperaturunterschiede) sowie Wasser und bestimmte Mineralstoffe.
Für die Lichtwahrnehmung sind v.a.zwei Pigmentsysteme, das Phytochromsystem und das Phototropinsystem verantwortlich. Das Phytochrom ist – wie das Chlorophyll und das Häm – ein Tetrapyrol, das allerdings keinen Porphyrinring bildet, sondern in offenkettiger Form vorliegt. Durch die Absorption von Photonen kann es seine Konfiguration ändern. Dadurch wirkt es wie ein lichtabhängiger Schalter. Bei Belichtung mit hellrotem Licht der Wellenlänge 665 nm wird Phytochrom in ein Molekül umgewandelt, das dunkelrotes Licht (735 nm) absorbiert, bei Belichtung mit dunkelrotem Licht wird dieses wieder in das Hellrot-absorbierende Phytochrom zurück verwandelt. Die jeweils zuletzt eingestrahlte Lichtqualität entscheidet über die ausgelöste Entwicklung. In der Zelle ist das Phytochrom an ein Protein gebunden. Das Phytochromsystem ist z. B. für die Samenkeimung von Licht- und Dunkelkeimern und für das extreme Streckungswachstum im Dunkeln (Etiolement, Vergeilung) verantwortlich.
Für die Krümmung von Pflanzensprossen zum Licht (Photropismus) ist nur der blaue Anteil des Lichts verantwortlich. Als wahrnehmendes Pigmentsystem konnte das Chromoprotein Phototropin nachgewiesen werden. Blaulicht wird von dem Apoprotein es Phototropins, einem Flavinmononucleotid, absorbiert und dies bewirkt, dass ein Phosphat an das Phototropin angehängt wird. Das so aktivierte Phototropin setzt eine Signaltransduktionskette in Gang, die dazu führt, dass im Sprossmeristem verstärkt das Phytohormon Auxin produziert wird. Es wird zunächst auf die dunkle Seite des Vegatationskegels verlagert und dann vorwiegend über die basalen Zellmembranen weitergegeben. Dafür sind bestimmte Effluxproteine und Afflux-Carrier verantworlich. Das Auxin verstärkt das Streckungswachstum der Zellen dadurch, dass die H+-ATP-aseaktivität und die K+-Aufnahme durch die Zellmembran verstärkt werden.
Abb. 10 Das Phytohormon Auxin verstarkt das Streckungswachstum von Zellen. Einseitig aufgetragene, Auxin-haltige Wuchsstoffpaste führt zum Krümmungswachstum eines Kalanchoe-tubilora-Sprosses.
Abb. 11 Phototropismus; Krümmung durch einseitige Auxinwirkung
Auch zur Wahrnehmung der Schwerkraft bzw. der Massenbeschleunigung haben Pflanzen sehr empfindliche Einrichtungen, die in mancher Hinsicht an Schweresinnesorgane von Tieren erinnern. Da in Schwerkraft-empfindlichen Pflanzenteilen wie Wurzelhauben, Keimscheiden und Sprossachsen auffällig assymmetrisch gelagerte Stärkekörner (in Amyloplasten) auftreten, spricht man auch von Statolithenstärke. Es wir vermutet, dass der entscheidende Reiz die partielle Dehnung der äußeren Zellmembran ist, die durch das Gewicht des Protoplasten ausgelöst wird. Die Statolithenstärke erhöht das Gewicht.
Viele Pflanzen – v.a. kletterne Sprossachsen und Ranken – können Berührungsreize wahrnehmen. Bei der fleichfessenen Venus-Fliegenfalle gibt es eine Reizwahrnehmung durch Sinnesborsten, ein Aktionspotenzial und eine Erregungsleitung . Noch ausgeprägter ist die Erregungsleitung bei der Sinnpflanze (Mimosa pudica).
Wurzelspitzen können chemische Reize wahrnehmen, z. B. Phosphationen, Wassermoleküle und Sauerstoffmoleküle. Auf diese Weise können Baumwurzeln kleinste Risse in unterirdischen Wasserleitungen aufspüren und in den Wasserleitungen dann verstopfende „Wurzelzöpfe“ bilden.
Sprachlos und gedankenlos?
Vor mehr als 35 Jahren erregte ein Buch mit dem Titel „Das geheime Leben der Pflanzen“ viel Aufsehen (Tompkins, Bird 1973). Die Autoren stellen darin „Pflanzen als Lebewesen mit Charakter und Seele und ihren Reaktionen in physischen und emotionalen Beziehungen zum Menschen“ dar. Sie schildern, wie eine ausgedehnte sprachliche Kommunikation mit Pflanzen möglich sei und wie man die Gefühle seines Gummibaumes oder seiner Palmlinie durch Zuspruch günstig oder ungünstig beeinflussen könne. Pflanzen werden in diesem Buch gewissermaßen als „bessere Menschen“ dargestellt. Von Botanikern und Pflanzenphysiologen wurde diese Veröffentlichung natürlich nicht ernst genommen und zu Recht ist es schnell still geworden um die angeblich objektiven Experimente der an Lügendetektoren und Oszilloskope angeschlossenen Pflanzen. Allerdings weiß man heute viel mehr über Signalaufnahme, Verarbeitung und Weitergabe von Pflanzen und über Formen pflanzlicher Kommunikation als vor 30 oder 40 Jahren und dabei wurden durchaus erstaunliche Fähigkeiten entdeckt. So besitzen Pflanzen ein großes Repertoire an Signalstoffen, die sie in Abhängigkeit von Umwelteinflüssen einsetzen können, um im eigenen Pflanzenkörper oder auch bei Nachbarpflanzen und sogar bei Tieren Reaktionen hervorzurufen.
Signalstoffe, die steuernd und regelnd in die inneren Entwicklungs- und Stoffwechselprozesse einer Pflanze eingreifen, werden in Analogie zu tierlichen Hormonen als Phytohormone bezeichnet. Phytohormone sind relativ kleine Moleküle, die Informationen von ihrem Bildungsort zu Zielzellen und Zielgeweben übertragen. In der Regel setzen sie in den Zielzellen eine Signaltransduktionskette in Gang, die zu einer Reaktion der Zelle führt . Diese Reaktion kann direkt im Cytoplasma stattfinden, sie kann aber auch über Transkriptionsfaktoren zum An- oder Abschalten von Genen führen. Dabei spielen Rezeptoren in der Zellmembran, verschiedene Proteine (G-Proteine, Proteinkinasen, phosphorylierte Proteine als Transkriptionsfaktoren) und kleine, als sog. sekundäre Botschafter (second messenger) wirkende Moleküle und Ionen wie Ca++, und cyclisches Adenin- bzw. Guanin-Monophosphat (cAMP, cGMP) eine Rolle (Tab. Phytohormone).
Viele Pflanzen produzieren chemische Abwehrstoffe, mit denen Fressfeinde oder Krankheitserreger abgewehrt werden können. Oft werden solche Stoffe erst produziert, wenn die Pflanzen von pflanzenfressenden Insekten oder anderen Herbivoren angeknabbert werden. Die Konzentrationen von Limonen und anderen Monoterpenen im Gewebe von Fichten und Lärchen zum Beispiel erhöhen sich als Reaktion auf Verletzungen. Limonen hat eine starke toxische Wirkung auf Borkenkäfer. Auch das Alkaloid Nikotin, das z. B. in Tabakpflanzen vorkommt, ist ein hochwirksamen Insektengift. In beschädigten Blättern erreicht es eine bis zu 10fach höhere Konzentration als in unbeschädigten.
Bestimmte durch Fraßfeinde hervorgerufene Verletzungen können Pflanzen dazu veranlassen, Stoffe wie Jasmonate als Signalstoffe an die Umgebung abzugeben. Diese Pheromone regen dann auch bei Nachbarpflanzen die Produktion von Abwehrstoffen an. Die Pheromonwirkung ist nicht nur auf Pflanzen derselben Art beschränkt. Manche Pflanzenarten setzen als Reaktion auf Herbivorenbefall sogar Substanzen frei, die die Parasiten dieser Pflanzenfresser anlocken. Maispflanzen, die durch den Fraß von Raupen der Zuckerrübeneule beschädigt wurden, produzierten unterschiedlich flüchtige Terpenoide, die nachweislich als Lockstoff für die auf den Raupen parasitierende Schlupfwespen (Cortesia marginiventris) wirken. . Die Signalketten, die von der Vielfalt flüchtiger Pflanzensubstanzen an ihren Zielen ausgelöst werden können, sind bisher zum großen Teil unbekannt, aber es gibt auch schon viele erforschte Beispiele (Abb. 10).
Besonders vielfältig ist der Signalaustausch der Pflanzen mit ihren Bestäubern, sowohl über optische als auch über chemische Signale
Abb. 12 Signalstoffe bei Pflanzen
Ebenso wie bei der tierlichen Kommunikation spielt auch bei Pflanzen die Täuschung eine Rolle (Kasten Duftmimikry bei Orchideen).
Interesse wecken für das Andere? – Pflanzenkunde unterrichten
Pflanzen sind für Kinder und Jugendliche i.d.R. weniger faszinierend als Tiere. Dies belegen mehrere empirische Studien (z. B. Hesse 2000). Während schon Kleinkinder von Tieren begeistert sind und ihnen ihre ganze Aufmerksamkeit zuwenden, sind Pflanzen selbst für Kinder im Grundschulalter in der Regel gar keine richtigen Lebewesen. Pflanzen, Nicht-Tiere, reagieren nicht, wenn man sie anspricht oder streichelt, sie bewegen sich nicht von der Stelle, sie bellen, brüllen oder zwitschern nicht und sie lassen keinen Schmerz erkennen. Die große Bedeutung, die Pflanzen für das Landschaftsbild haben, ist für Kinder ebenfalls nicht wichtig, da sich Interesse und Gefühle für Landschaftsästhetik erst später entwickeln. Allerdings gibt es durchaus animierende Gefühle zu Pflanzen, die mit Ästhetik, Abenteuerlust oder Freude an technischen Funktionen zu tun haben:
Aus Pflanzen etwas gestalten: Blumenstrauß-Pflücken, Kränze und Blumenketten binden…
Aus Pflanzenteilen etwas konstruieren: Weidenpfeifen und Panflöten bauen, Wasser durch Löwenzahnstängel leiten, aus einem Schilfblatt ein Segelschiffchen bauen, einen Haselzweig zum Flitzebogen machen…
Pflanzenumgebungen für Abenteuer nutzen: auf Bäume klettern und Baumhäuser bauen, an einer Liane schaukeln, Baumstämme als Wippe nutzen, im Maisfeld Verstecken spielen, sich in Laubhaufen vergraben…
Auch das Phänomen der Samenkeimung und des Wachstums oder das Ableger Großziehen kann Kinder und Jugendliche faszinieren.
Pflanzenkunde-Unterricht muss an solche Erfahrungen und Vorstellungen anknüpfen. Entsprechende Versuche hat es in der Biologiedidaktik immer wieder gegeben: UB 184 „Kreative Botanik“, UB 275 „Pflanzen züchten und vermehren“, UB 286 „Außergewöhnliche Pflanzen“, „Pflanzen stellen sich vor“, „Gärten zum Leben und Lernen“ usw.
Im Rahmen des Modellversuches „Praxis integrierter naturwissenschaftlicher Grundbildung“ (PING) des Landes Schleswig-Holstein wurde für den 5/6. Jahrgang eine UE „Ich und die Pflanzen“ entwickelt, bei der der lebensweltliche Bezug der einzelnen Themen besonders wichtig genommen wird. Dabei wird allerdings in Kauf genommen, dass es oft nicht eigentlich um „Pflanzenkunde“ sondern um Selbsterfahrung („Phantasiereise zur blauen Blume“) oder um irgendwelche Nutzungen von Pflanzen oder pflanzlichen Produkten geht („Wie energiesparend kann ich mit Holz kochen“). Dabei ist der Ansatz sicherlich richtig, bei der Pflanzenkunde verstärkt von eigenen Erfahrungen oder Erfahrungen mit Tieren auszugehen, z. B. durch Fragestellungen wie „Können Pflanzen sehen?“, „Können Pflanzen atmen?“, „Können Pflanzen fühlen?“, „Können Pflanzen laufen, schwimmen, fliegen, klettern…?“ oder „Können Pflanzen um Hilfe rufen?“ . Dabei könnte von der eigenen Erfahrung (Wie klettere ich? Welche Hilfsmittel nutze ich zum Klettern?) die Funktion des Kletterns bei Pflanzen und dann der Bau (Pflanzenlösung des Funktionsproblems) behandelt werden. Ästhetische Erlebnisse mit schönen Blüten oder bizarren Blattmustern können der Ausgangspunkt für Fragen nach der biologischen Funktion und der physikalischen Grundlage der Phänomene sein – z. B. leuchtende Blütenfarben durch Totalreflexion an Interzellularen, Fettglanz durch Reflexion an Stärkeschichten, Samteffekt durch Lichtabsorbtion an feinen Papillen usw.
Gerade weil Kinder Pflanzen zunächst als keine echten Lebewesen oder Mitgeschöpfe ansehen, ist der Überraschungseffekt groß, wenn sie an Pflanzen tierliche Fähigkeiten entdecken können, z. B. rasche Bewegungsreaktionen auf Reize oder das „Fleischfressen“. Dabei kann sich die Lehrkraft Charles Darwin zum Vorbild nehmen. In seiner Autobiografie schreibt er: „Es hat mir immer große Freude bereitet, die Pflanzen in der Stufenleiter organisierter Wesen zu erhöhen. Ich empfand daher ein besonderes Vergnügen, als ich zeigen konnte, wie viele und wie wunderbar schön angepasste Bewegungen die Spitze einer Wurzel besitzt“ (Darwin, Erinnerungen…1876-1881, Aulis,Köln 1982, S.159).
Für unmittelbare Erfahrungen, Beobachtungen und Experimente am lebenden Objekt sind Pflanzen besonders gut geeignet., da die Untersuchungsobjekte i. d. R. leicht zu beschaffen sind , und auch, weil ethische Probleme, wie sie beim Experimentieren mit Tieren auftreten können, hier keine Rolle spielen. Schließlich stehen Pflanzen beim „Biologieunterricht im Freien“, bei Geländepraktika, Exkursionen, Schulgartenprojekten und Klassenfahrten schon deshalb im Vordergrund, weil sie die Struktur einer Lebensgemeinschaft maßgebend prägen, weil sie als leicht zugängliche Zeigerorganismen dienen können und weil man über Pflanzenarten viele ökologische Zusammenhänge erschließen kann. Auch wenn bei der Freilandbiologie ökologische Fragestellungen im Vordergrund stehen, kann man gerade auf Exkursionen auch viele Form- und Funktionszusammenhänge beobachten, erleben und verstehen.
Der richtige Weg einer zwar „humanzentrierten Pflanzenkunde“, der aber dazu führt, dass die Lernenden auch die Pflanze selbst und nicht nur ihre Beziehung zu derselben als interessant begreifen, bleibt das schwer erreichbare aber lohnende Ziel.
Quellen
Adl, S., M. u. a.( 2005) :The New Higher Level Classification of Eukaryotes with Emphasis on the Taxonomy of Protists. J. Eukaryot. Microbiol., 52(5), 2005 pp. 399–451 http://myweb.dal.ca/asimpso2/sopclass.htm
Bell, A.D. (1994): Illustrierte Morphologie der Blütenpflanzen, 335 S.. Stuttgart: Ulmer
Bell, P.R., Hemsley, A.R. (20002): Green plants. Their origin and diversity. Cambridge Univ. Press
Hesse, M. (2000): Erinnerungen an die Schulzeit – Ein Rückblick auf den erlebten Biologieunterricht junger Erwachsener. Zeitschrift für Didaktik der Naturwissenschaften 6, 187-201
Institut für Qualitätssicherung Schleswig-Holstein (IQSH): PING–Material (2006):
Probst, W. (1999): Halme und Stängel. In: Deutsche Gartenbaugesellschaft/G. Hütten (Hrsg.): Wege zur Naturerziehung, S. 116-133
Probst, W., Hrsg.(1998-2001): Gärten zum Leben und Lernen. Seelze: Kallmeyer (16 Hefte)
Probst, W. (20072): Pflanzen stellen sich vor. Köln: Aulis
Probst, W. (2009): Stoffkreisläufe. Basisartikel in UB 349 (33.Jg), S.2-11. Seelze: Friedrich
Probst, W., Schuchardt,P., Hrsg. (20205): Basiswissen Biologie, Abitur. Berlin: BI-Duden
Purves, W. K. u. a. (201910): Biologie. München: Elsevier
Kadereit, J. W. u.a.(201437): Lehrbuch der Botanik (Ersthrsg. E. Strasburger) Heidelberg: Spektrum
Seyffert, W.: Lehrbuch der Genetik. Spektrum, Heidelberg/Berlin 20032
Taiz, L., Zeiger, L., Møller, I., M., Murphy, A. (2018): Plant Physiology and Development. Oxford: Sinauer
Urry, L. A. u.a. (201911): Campbell Biologie. München:. Pearson Deutschland
Whittaker, R. (1969): New concepts of kingdoms or organisms. Evolutionary relations are better represented by new classifications than by the traditional two kingdoms.. Science163: pp. 150–160 doi:10.1126/science.163.3863.150. PMID5762760.
Dieser Beitrag beruht auf Recherchen, die ich im Zusammenhang mit dem Unterricht Biologie Heft „Naturschutz auf neuen Wegen“ (UB 465) durchgeführt habe. Das Heft ist im Sommer 2021 erschienen.
Seit Beginn der Industrialisierung haben sich die Verhältnisse auf unserem Bioplaneten Erde (Kattmann 1991,2004) durch exponentielles Wachstum von Wirtschaft und Bevölkerung drastisch verändert, besonders deutlich in den letzten Jahrzehnten. Dank der elektronischen Datenverarbeitung und immer genaueren Registrierungsmöglichkeiten durch Satelliten lassen sich diese Veränderungen recht genau beschreiben. Schon lange vorher gesagt aber erst in den letzten Jahren in den Mittelpunkt des kollektiven Bewusstseins gerückt ist die durch menschliche Aktivitäten verursachte Klimaerwärmung, um die Dimension dieser drastischen Entwicklung besonders zu betonen, wird neuerdings von „Klimaerhitzung“ gesprochen. Obwohl diese negativen Veränderungen besorgniserregend rasch voranschreiten, besteht nach wie vor Hoffnung auf eine Stabilisierung. Es gibt viele Ideen und auch schon realisierte Beispiele, wie man die Zukunft des Bioplaneten nachhaltiger gestalten könnte.
Bioplanetenschutz heißt Schutz der Funktionsabläufe
Nach konservativen Verständnis geht es im Naturschutz um den Erhalt oder gegebenenfalls auch die Wiederherstellung eines jetzigen oder früheren Zustandes, der den Menschen und seine Aktivitäten weitgehend ausklammert. In einem erweiterten Verständnis bedeutet der Schutz der Natur Schutz des Bioplaneten, d. h. insbesondere Schutz und Erhalt der Funktionsabläufe. In diesem Sinne können auch weitgehende Eingriffe und Manipulationen durch den Menschen (Geoengineering, synthetische Biologie), ökonomisch Maßnahmen wie Steuererhebungen oder juristische Maßnahmen wie Verbote von Verbrennungsmotoren oder Kohlekraftwerken als Naturschutzmaßnahmen verstanden werden.
Für die Rechtfertigung solcher Eingriffe sind einmal auf breiter wissenschaftlicher Basis erstellte Analysen und Prognosen erforderlich. Zum anderen müssen diese Erkenntnisse Grundlage von Bildung und Ausbildung werden. Neben neuen technischen Lösungen muss Naturschutz deshalb verstärkt um die menschliche Akteure einschließen. Sozio-ökonomische Aspekte müssen mit gedacht und interdisziplinär behandelt werden. Dazu gehören besondere Anreize für umweltfreundliches oder naturschutzkonformes Verhalten, deren Vorteile unmittelbar wirksam werden. Nur dann wird es möglich sein, den demokratischen Konsens herzustellen, der für eine politische Durchsetzung sinnvoller Maßnahmen notwendig ist.
Landschaftsgestalterische Maßnahmen können zur Renaturierung oder sogar Regenerierung von Ökosystemen führen oder neue artenreiche Ökosysteme entstehen lassen.
Die Wiedervernässung von Mooren kann deren Fähigkeit wieder herstellen, Kohlenstoff in unvollständig abgebautem Pflanzenmaterial zu speichern. Außerdem wirken die Torfkörper der Moore regulierend auf den Wasserhaushalt.
Die naturnahe Gestaltung von stillgelegten Kiesgruben, Steinbrüchen und Tagebauflächen (z. B. Braunkohle) kann ökologisch wertvolle Biotope und Landschaften entstehen lassen und damit die Biodiversität fördern.
Entrohrung, Renaturierung und Remäandrierung von Bachläufen kann die Wasserqualität verbessern, Überschwemmungsgefahren mindern und im Sinne eines natürlichen Wasserkreislauf wirken. Außerdem entstehen dadurch vielseitige Lebensräume, welche die Biodiversität fördern.
Die Anlage von marinen Hartsubstratböden, z. B. um Offshore-Windparks kann die Biodiversität fördern, insbesondere durch die Schaffung neuer Siedlungsflächen für Aufwuchsorganismen und Brutgebiete für Fische.
Durch geeignete Maßnahmen können bisher eher als Plantagen genutzte Waldgebiete in naturnahe Wälder umgebaut werden.
In potenziellen Waldgebieten kann der Anteil der Bewaldung durch Aufforstungsmaßnahmen erhöht werden.
Vor allem in Trockengebieten können überweidete Landschaften durch Regulierung des Weidegangs aufgewertet werden.
Durch Überweidung desertifizierte Landschaft in Nordafghanistan bei Kunduz,25.7.1974 (Foto W.Probst)
Für diese Renaturierungs- und Regenerationsmaßnahmen werden viele Arbeitskräfte benötigt. Durch entsprechende Förderprogramme können Landwirtschaft und Forstwirtschaft in Renaturierungsprogramme eingebunden werden.
Eine weitere Möglichkeit bestünde darin, für solche Aufgaben verstärkt das Militär einzusetzen und dafür entsprechende Kenntnisse und Fertigkeiten in die militärische Ausbildung einzubauen (J. Ellington in Randers 2012).
Besonders spektakuläre Großprojekte sind Chinas „Grüne Mauer“ und die 2005 diesem Vorbild folgende von der Afrikanischen Union initiierte grüne Mauer durch die Sahelzone . Sie sollen Wüstenbildung aufhalten und teilweise rückgängig machen.
Die chinesische „Grüne Mauer“ verdankt ihren Namen der chinesischen „Großen Mauer“: Während die Große Mauer Schutz gegen die Völker aus dem Norden bieten sollte, soll die Grüne Mauer vor Wüstenstürmen schützen. Das Projekt wurde schon 1978 begonnen und soll bis 2050 fortgesetzt werden. Bis dahin sollen 350.000 km² – dies entspricht etwa der Fläche der Bundesrepublik – mit Bäumen bepflanzt sein. Dabei besteht allerdings die Gefahr, dass durch die Bewässerung der neu angelegten Schutzwälder alte, flussbegleitende Wälder geschädigt werden (Missall u.a. 2018).
Afrikas „Grüne Mauer“ (GGWSSI; Great Green Wall of the Sahara and the Sahel Initiative) ist als 7775 km langer, mindestens 15 km breiter Baumstreifen geplant, der die Trockenregion am südlichen Rand der Sahara von Dakar bis Dschibuti durchziehen soll. Die Idee geht auf den 1987 ermordeten Präsidenten von Burkina Faso Thomas Sankara und auf die kenianische Professorin und Nobelpreisträgerin Wangari Maathai und ihr „green belt movement“ zurück. Unter der Präsidentschaft des damaligen Präsidenten von Nigeria Olusegun Obasanjo übernahm die Afrikanische Union das Projekt. Bisher wird es von 22 afrikanischen Staaten unterstützt. Mittlerweile sprechen viele Verantwortlichen nicht mehr von einer Mauer sondern eher von einem Mosaik, da verstärkt in Dorfgemeinschaften verwurzelte Projekte unterstützt werden sollen. Außerdem soll auch der Erhalt und Schutz bereits existierender Baumbestände stärker gefördert werden. Auf dem „One Planet Summit“ im Januar 2021 in Paris hat die internationale Gemeinschaft 11,8 Mrd. Euro für das Projekt zugesagt.
Über diese und zahlreiche weitere Aufforstungsprojekte berichtet Daniel Schilk in seinem 2019 erschienenen Buch „Die Wiederbegrünung der Welt“.
Ökosystemerhalt durch assistierte Evolution
Die Idee, gefährdete Arten dadurch zu erhalten, dass man sie in Gefangenschaft oder im Labor züchtet und dann in natürlichen Ökosystemen freilässt, ist schon mehr als 100 Jahre alt.1895 hat der Geschäftsmann und Ornithologe Edward McIlhenny auf diese Weise in Louisiana die vom Aussterben bedrohten Schmuckreiher erhalten. Zwischen 1885 und 1807 konnte Richard Henry den neuseeländischen Kakapo (flugunfähiger Papagei) und den Kiwi durch Translokation von Tieren auf die vor der Westküste Neuseelands liegenden Insel Resolution Island vor dem Aussterben retten (Seddon 2017). Mittlerweile gibt es viele mehr oder weniger erfolgreiche Beispiele solcher Versuche, durch Translokation oder Zucht und Aussetzen gefährdete Arten zu erhalten, in Mitteleuropa zum Beispiel Luchse, Biber und Waldtrappe. Dabei geht es nicht nur um den Erhalt der betreffenden Arten sondern auch um die Funktion der Ökosysteme. Durch die Wiederetablierung von Schlüsselarten hofft man, Ökosysteme zu regenerieren oder auch neue wertvolle Ökosysteme zu schaffen.
Doch auch über weitergehende Schritte wird nachgedacht. Dabei könnte die synthetischen Biologie eine wichtige Rolle spielen, indem ausgestorbene Arten wie das Wollhaar-Mammut oder der Auerochse gentechnisch rekonstruiert werden (De-Extinction, Redford 2017). Als Quelle könnte genetisches Material aus alten Sammlungen oder aus Fossilien und verwandte noch lebende Arten genutzt werden.
Die Überlegungen gehen noch einen Schritt weiter: Es können nicht nur natürliche Arten künstlich vermehrt oder wiederhergestellt, sondern auch „verbessert“, also durch Zucht oder Gentechnik gezielt verändert werden. Bei Riffkorallen soll zum Beispiel versucht werden die endosymbiontisch Zooxanthellen gentechnisch so zu verändern, dass sie auch bei höheren Meerestemperaturen funktionsfähig bleiben und dadurch Korallenbleiche vermieden werden können. Allgemein soll es durch das Einbringen solcher „verbesserter“ Lebewesen, die veränderte Umweltbedingungen besser aushalten,gelingen Ökosysteme als Ganzes zu erhalten.
Erhalt, Regeneration und Neuschaffung von Ökosystemen mit Hilfe Assistierter Evolution (Grafik W.Probst)
Verhinderung der Klimaerwärmung durch Geoengineering
Durch technische Eingriffe in das Klimasystem (Geoengineering) soll die Klimaerwärmung vermindert werden. Dabei sind vor allem zwei Möglichkeiten denkbar:
Der Atmosphäre werden direkt Treibhausgase, insbesondere Kohlenstoffdioxid, entzogen (Carbon Dioxid Removal CDR, Carbon Capture and Storage, CCS).
Die auf die Erde eintreffende Sonnenstrahlung wird verringert (Solar Radiation Management SRM).
Methoden des Geoengeneering (W. Probst verändert nach Angaben in Gynsky u.a. 2011)
Die Bindung von Kohlenstoffdioxid kann entweder terrestrisch oder marin erfolgen. Klassische Vorschläge beruhen auf Methoden, durch die der Aufbau von Biomasse – zum Beispiel durch großflächige Aufforstung – gefördert wird oder Kohlenstoff haltiges Material in den Boden eingearbeitet wird (Beispiel Terra Preta). Auch Möglichkeiten, CO2 direkt aus der Luft zu filtern und unterirdisch dauerhaft zu speichern – zum Beispiel durch Einpressen in tiefliegende geologische Formationen (Carbon Capture and Storage, CCS). Die meisten derzeit laufenden Pilotprojekte testen die Integration dieser Art der CO2 Abscheidung direkt in der Kombination mit Kohlekraftwerken, weil dort in den Abgasen der CO2 Gehalt hoch ist. Die Möglichkeit der direkten Filterung aus der Luft, in der CO2 derzeit höchstens zu 0,5 Volumenpromille enthalten ist, wäre bisher zwar möglich aber sehr kostenaufwendig.
Um CO2 verstärkt in den Ozeanen zu binden, wird die Ozeandüngung diskutiert. Dabei bedient man sich der sogenannten biologischen Pumpe. Kohlenstoffdioxid wird von Mikroalgen assimilert und ein Teil davon wird als dauerhaftes Kohlenstoff-haltiges Sediment am Meeresboden abgelagert. Durch Düngung könnte die Phytoplanktonproduktion angeregt werden. Da man von den Makronährmineralien Nitrat und Phosphat sehr große Mengen benötigen würde, hat man bei bisherigen Versuchen mit dem Mikronährmineral Eisen gearbeitet Entsprechende verhältnismäßig kleinräumige, zeitlich begrenzte Versuche, die zu Beginn des Jahrhundert durchgeführt wurden, hatten allerdings wenig überzeugende Ergebnisse. Zwar konnte man zunächst Algenblüten bewirken, aber das Absinken des Phytoplanktons trat nur in sehr geringem Maße ein. Ein großer Teil wurde vom Zooplankton aufgenommen und dadurch veränderten sich die Nahrungsnetze. Auch die Blüte von toxischen Kieselalgen konnte beobachtet werden. Zudem ist die kontinuierliche Düngung sehr energieaufwendig und die Bilanz des tatsächlich gebundenen CO2 ist dadurch viel geringer als zunächst theoretisch berechnet wurde.
Eine weitere Möglichkeit, die Phytoplanktonproduktion zu erhöhen, läge in der Manipulation der marinen Schichtung. Wenn man verstärkt nährmineralreiches Tiefenwasser in obere Wasserschichten verlagern könnte – wie dies unter derzeit natürlichen Bedingungen zum Beispiel an der Westküste des amerikanischen Kontinents geschieht – könnte man die Phytoplanktonproduktion anregen. Entsprechende aus langen Rohren bestehende Pumpen, die vom Wellenschlag angetrieben werden, wurden zwar erfolgreich konstruiert. Um einen messbaren Effekt bei der marinen CO2– Speicherung zu erreichen, wären allerdings eine sehr große Zahl solcher Pumpen notwendig und die Folgewirkungen sind schwer abzuschätzen.
Außer durch die biologische Pumpe wird auch durch eine physikalische Pumpe CO2 von der Oberfläche in die Tiefen der Weltmeere befördert. Kalte Wassermassen mit hohem Salzgehalt im Nordatlantik und in dem antarktischen Zirkularstrom sinken ab und setzen globale Meeresströmungen in Gang, bei denen es an anderer Stelle zum aufsteigen von Tiefenwasser kommt. Da CO2 in kaltem Wasser eine höhere Löslichkeit hat als in wärmeren Wasser, wird durch diesen Prozess langfristig CO2 aus der Atmosphäre in die tieferen Wasserschichten transportiert. Aber alle Methoden, die bisher versucht wurden, um diesen Absinkeprozess zu verstärken, waren nicht erfolgreich, insbesondere, weil das Absinken des Wassers an anderen Stellen den Auftrieb verstärken und damit kohlenstoffdioxidreiches Wasser an die Oberfläche befördern würde. Ob die Bilanz dann tatsächlich zu einer verstärkten marinen CO2– bzw. C-Speicherung führen würde, ist fraglich.
Die zweite Möglichkeit ist die Verringerung der auf der Erde auftretenden Sonnenstrahlung, also die Beeinflussung des Strahlungshaushaltes (Solar Radiation Management SRM). Sie beruht einmal auf Methoden, welche die Reflexion der Strahlung verstärken, also die Erhöhung des Albedos der Erdoberfläche. Diskutiert wird zum Beispiel das Weißeln von Dachflächen oder die Installation von großen Reflektorflächen in Wüsten oder auf Meeren. Zur zum anderen könnte das Einbringen von Aerosolen in die Stratosphäre oder von großflächigen Spiegeln in den Weltraum das Durchdringen der Sonnenstrahlen bis zur Erdoberfläche verringern. Alle diese Methoden sind höchst umstritten, da man nur schwer Aussagen über die dabei auftretenden Nebeneffekte und Folgen machen kann. Außerdem ist der finanzielle Aufwand sehr hoch.
Insgesamt birgt Geoengineering große Risiken. Wenn sich aber zeigt, dass die vom Weltklimarat 2018 festgelegten Klimaziele anders nicht erreicht werden können, wird man die Risiken einiger solcher Methoden wahrscheinlich in Kauf nehmen (Ginsky u.a. 2011).
Kreislaufwirtschaft zur Abfallvermeidung
Vermeidung von Abfall und Umweltverschmutzung muss nicht (nur) auf Sparsamkeit und Verzicht aufgebaut sein, mindestens genauso wichtig ist eine konsequente Kreislaufwirtschaft: Alle Produkte müssen so konzipiert und hergestellt werden, dass sie „rematerialisierbar“ sind, ob Möbel, Kleider, Autos, Baumaschinen Häuser oder Lebensmittelverpackungen. Nach Ansicht des Chemiker und Designers Michael Braungart und des Architekten William McDonough ist dieses „cradle to cradle-Prinzip“ (C2C, „Von der Wiege zur Wiege“) sogar alleine entscheidend. (McDounough, Braungart 2009). Sie berufen sich dabei auf die Natur als Vorbild. Die üppigsten und artenreichsten Ökosysteme, die tropischen Regenwälder, sind nicht nur die produktivsten, sie setzen auch die größten Stoffmengen um. Daraus folgert Braungart, dass es nicht darum gehen kann, zu „sparen“ also, weniger umzusetzen, sondern darum, nicht zu „verbrauchen“ sondern zu „gebrauchen“. „Verschwendet! Aber richtig: Macht keinen Müll!“ fordert er. Sonnenenergie steht im Prinzip soviel zur Verfügung, dass es kein Problem ist, verschwenderisch damit umzugehen. Soziale Ungerechtigkeit und das Nord-Süd-Ungleichgewicht können nicht durch Sparsamkeit gelöst werden. Ihre Lösung ist aber Voraussetzung für geordnete, friedliche Verhältnisse auf unserem Planeten.
Dieses Konzept steht in gewissem Widerspruch zu der Forderung einer verminderten Ressourcennutzung wie sie vom Wuppertal Institut für Klima,Umwelt, Energie, zunächst als „Faktor 4“ (v. Weizsäcker, Lovins, Lovins 1995) später als „Faktor 10“ (Schmidt-Bleek 1997) propagiert wurde. Sicher kann es bei einer zukunftsfähigen, nachhaltigen Wirtschaft nur um ein „Sowohl-als-auch“ gehen, denn Kreislaufprozesse ganz ohne Abfall und Umweltschäden – das zeigt auch das Vorbild Natur – gibt es nicht. Fossile Brennstoffe sind ein Beispiel für solche natürlichen Abfälle und globale Katastrophen. Gutes Beispiel für die menschliche Wirtschaft ist die große Verschwendung von Nahrungsmitteln und die damit verbundene Zerstörung von gut funktionierenden Kreislauf-Ökosystemen und inhumaner Nutztierhaltung.
Wie zukünftiges Wirtschaften verbessert werden könnte zeigt ein in Dänemark entwickelter Industriepark, in dem eine „Symbiose“ zwischen verschiedenen Industrieunternehmen nicht nur eine starke Abfallverminderung sondern auch eine bessere Energienutzung ermöglichen (Kalundborg Symbiosis 2020).
Das größte Problem beim Plastikabfall sind die Verpackungen. Eine konsequente Einführung von kompostiertem Verpackungsmaterial könnte hier große Verbesserungen bringen. Weltweit hat die sehr erfolgreiche Einführung von Kaffeepads aus Kunststoff oder Aluminium zu einem enormen Anstieg von Verpackungsmüll und Ressourcenverbrauch geführt, jährlich mittlerweile über 40 Milliarden Kapseln. Aber immer mehr Firmen versuchen, kompostierbare Verpackugen zu produzieren. Ein Beispiel ist die Firma Nexe Innovations, die derzeit mit ihren kompostierbaren Kaffeepads recht erfolgreich ist, die in allen gängigen Kaffeemascinen verwendet werden können.
Neobiota-Management
Im Laufe der Erdgeschichte zerbrachen Kontinente oder schoben sich zusammen, Inseln und Inselarchipele entstanden neu oder gingen unter, aus Grabenbrüchen wurden Ozeane, Meeresbuchten wurden abgetrennt, Binnenmeere öffneten sich zum Ozean. Diese geologischen Ereignisse wurden begleitet von Ausbreitung, Rückgang, Einwanderung und Auswanderung von Lebewesen. Die Invasion neuer Arten und die Ausbreitung von Krankheitserregern und die dadurch bedingten Veränderungen von Ökosystemen sind ein natürlicher Vorgang in der Geschichte des Lebens. Doch im Gegensatz zu den geologischen Veränderungen haben die anthropogen verursachten globalen Veränderungen der letzten Jahrhunderte und vor allem der letzten Jahrzehnte zu einer enormen Beschleunigung dieser Invasionen beigetragen.
Schon im Zeitalter der europäischen Eroberungen und Kolonisationen und der Einwanderung von Europäern nach Amerika und Australien wurden Tier- und Pflanzenarten von Menschen gezielt von Kontinent zu Kontinent verbreitet.
In den letzten Jahrzehnten haben der globale Warenaustausch und der Reiseverkehr, aber auch die gezielte Einfuhr gebietsfremder Arten, zu einer starken Zunahme von Neobiota (Neubürgern) geführt. Diese Einwanderer sind ein ernst zu nehmendes Naturschutzproblem geworden. Durch die Verdrängung einheimischer Arten können sie Ökosysteme verändern und schließlich das Aussterben von Arten bewirken („invasive Arten“). In der EU-Liste invasiver gebietsfremder Tier- und Pflanzenarten („Unionsliste“) werden derzeit 66 Tier- und Pflanzenarten als möglicherweise invasiv aufgelistet. Bereits in Deutschland etabliert sind zum Beispiel der Riesen-Bärenklau (Heracleum mantegazzianum), das Indische Springkraut (Impatiens glandulifera), der Kamberkrebs (Orconectes limosus) und die Amurgrundel (Percottus glenii) (NABU 2019). Neben einer Konkurrenz mit einheimischen Arten geht es dabei auch um Schädlinge wie Kartoffelkäfer, Asiatischem Marienkäfer, Varoamilbe oder Buchsbaumzünsler, gegen die ansässige Arten kaum Abwehrkräfte entwickelt haben.
Wegsaum mit Drüsigem Springkraut (Impatiens glandulifera) im Rotwildpark Stuttgart, September 1991. Die Art stammt aus dem Himalaja und wurde 1839 nach England eingeführt. Von dort gelangte sie auf den Kontinent. Heute gilt sie als invasiver Neophyt und wird teilweise bekämpft. Verschiedene Untersuchungen zeigen jedoch, dass die Pflanze die natürliche Waldverjüngung kaum negativ beeinflusst (Foto W. Probst).
Besonders gefährdet durch invasive Arten waren und sind Inseln mit speziellen Ökosystemen und vielen endemischen Arten. Die absichtliche Aussetzung von Ziegen und Schweinen und die unabsichtliche Einfuhr von Ratten durch die frühen Seefahrer des 16.-19. Jahrhunderts hatten schon verheerende Auswirkungen auf pazifischen Inseln, aber auch die Besiedlung von Amerika, Australien und Neuseeland durch Europäer hat einen gewaltigen Invasionsschub verursacht, der das Ende zahlreicher einheimischer Arten bewirkte. Gut dokumentiert ist der Artenrückgang auf der Pazifikinsel Guam, der durch die eingeschleppte Braune Nachtbaumnatter (Bioga irregularis) verursacht wurde (Probst 2010).
Aber sind alle Neobiota problematisch? Einer der führenden Neobiota-Forscher, Ingo Kowarik, gibt darauf folgende Antwort:
Ja, wenn Veränderungen von Natur als Problem gesehen werden.
Ja wenn „Fremdes“ als negativ gesehen wird.
Nein, wenn unterschiedliche Auswirkungen berücksichtigt werden.
(Ingo Kowarik bei einem Vortrag zum Landesbiologentag an der Universität Hohenheim am 7.11.2020).
Durch auf wissenschaftlichen Grundlagen erarbeitete Management-Pläne versucht man, schädliche Auswirkungen von Neobiota auf die Biodiversität zu begrenzen. Ein Beispiel: Durch den organsierten Austausch von Ballastwasser in der marinen Schifffahrt seit 2017 soll die Einschleppung gebietsfremder Arten verhindert werden.
Pandemien und Naturschutz
Mit dem globalisierten Austausch von Menschen und Waren haben sich auch Krankheitserreger ausgebreitet. Dies führte nicht selten in den neuen Ausbreitungsgebieten zu verheerenden Epidemien. Besonders betroffen waren indigene Bevölkerungsgruppen Amerikas, zum Beispiel die mittlerweile (fast?) ausgestorbenen Ureinwohner Feuerlands, die Yagan oder Yamana (Kaiser 2013).
Auch in umgekehrter Richtung wurden schon lange Keime übertragen, zum Beispiel der Cholera-Erreger Vibrio cholerae aus Indien. Auch die Übertragung von Krankheitserregern von Tieren auf Menschen geht bis in das Neolithikum zurück, als durch die Einführung der Nutztierhaltung der Kontakt zwischen Tieren und Menschen enger wurde. Masern und Tuberkulose stammen von Kühen, Keuchhusten von Schweinen und Grippe von Enten (Shah 2020).
Die rasant voranschreitende Globalisierung der letzten Jahrzehnte hat die rasche Ausbreitung von Krankheitserregern, insbesondere von Bakterien und Viren, weiter gefördert. Dabei spielen nicht nur die größere Mobilität der Bevölkerung und der Reiseverkehr über große Entfernungen eine wichtige Rolle, sondern auch die immer stärkere Einschränkung von Wildtierpopulationen durch Verlust natürlicher Lebensräume, zum Beispiel tropischer Regenwälder. In den kleineren Populationen können sich Erreger schneller ausbreiten. Außerdem fördert der immer intensivere Kontakt der ständig wachsenden menschlichen Bevölkerung mit Tieren früher sehr abgelegener Regionen den Übergang von Krankheitskeimen von Wildtieren zu Menschen (Beispiel AIDS, Ebola, Vogelgrippe H1N5, SARS-Corona, Covid 19; vgl. Ruppert 2021, Keesing 2010, Jones 2008).
Man kann nur hoffen, dass die derzeitigen Erfahrungen mit der Covid 19 Pandemie zu einem Umdenken und einer vorsichtigeren Vorgehensweise führen.
Die immer intensivere Einflussnahme des Menschen auf alle Lebensräume und die räumliche Einschränkung naturnaher Biotope sollte gestoppt und womöglich rückgängig gemacht werden. Dabei geht es insbesondere darum, die Vielfalt der Arten in ausreichender Populationsgröße zu erhalten. Dadurch kann erreicht werden, dass sich Viren, auch neue mutierte Viren, nicht flächendeckend ausbreiten, sondern eher in einer Nische bleiben und nach einiger Zeit wieder Aussterben (infektionsbiologischer Verdünnungseffekt). Auch Generalisten wie Ratten oder Sperlinge, die für die Übertragung auf menschliche Populationen besonders gefährlich sind, sind in intakten Ökosystemen weniger verbreitet .
Inklusiver Naturschutz
Naturschutz sollte nicht nur in abgegrenzten Gebieten oder Biotopen stattfinden sondern überall. Die Einrichtung von Naturschutzgebieten hat zwar insofern eine gewisse Berechtigung, als es leichter ist, ökologisch wertvolle Lebensgemeinschaften, Schlüsselarten und Habitate auf diese Weise zu schützen. Außerdem sind naturnahe, von Menschen wenig beeinflusste Gebiete eine wichtige Voraussetzung für die ökologischen Funktionen des Bioplaneten. Es besteht aber die Gefahr, dass außerhalb von Schutzgebieten auf Natur und natürliche Funktionsabläufe keine oder zu wenig Rücksicht genommen wird. Angesichts der immer intensiveren Nutzung der Erde durch den Menschen wird es außerdem immer schwieriger, ausreichende Flächen für ungenutzte Gebiete bereitzuhalten. Flächendeckender „inklusiver“ Schutz der Natur auch in Städten und Gewerbegebieten, in Agrarlandschaften und entlang von Verkehrswegen wird deshalb immer wichtiger. Es gibt mittlerweile viele Ansätze, wie Natur auch außerhalb von Schutzgebieten nicht „ausgeschaltet, sondern eingeschaltet“ werden kann (Le Roy 1973), und Biodiversität und natürliche Funktionsabläufe erhalten bleiben.
Städte und Siedlungen
Zwischen 1985 und 2015 hat die die Ausdehnung von Städten und Siedlungen jährlich um 9687 km² zugenommen, mit steigender Tendenz (Liu et al. 2020). Damit ist der Flächenverbrauch der Städte schneller gewachsen als die Bevölkerung. Für eine nachhaltige Entwicklung müssen Städte deshalb „ökologisch“ werden. Eine Stadt mit großen Grünanlagen wie Parks und Gärten bietet zwar eine hohe Lebensqualität und eine bessere Ökobilanz. Dies geht aber insofern auf Kosten der Umgebung, als sie mehr Fläche für denselben umbauten Raum benötigt. Eine Erfolg versprechende Möglichkeit für dicht bebaute Großstädte ist die Integration von Bauwerken und Grünanlagen.
Neben Minderung des Klimawandels durch eine Verbesserung der CO2-Bilanz können dadurch auch die Auswirkungen einer Klimaerwärmung verringert werden (Grewe 2020). Schließlich wirken mit Sachverstand begrünte Städte auch dem Verlust der Biodiversität entgegen.
Die dynamische Vergrößerung städtischer Flächen von1985-2015. Datengrundlage sind Landsataufnahmen mit einer Auflösung von 30m. b) Steigungsrate des Stadtflächen-Wachstums auf den verschiedenen Kontinenten (Quelle Liu et al. 2020).Vernetzte Dachgärten (Zeichnung W.Probst)
Dächer
Schon lange zählt es zu Attributen ökologischer Bauweise, Dächer zu begrünen. Die Etablierung und Ausgestaltung solcher Dachgärten und Wiesen ist aber noch sehr stark ausbaufähig, wie man auf Luftbildern von Städten leicht erkennen kann. Begrünte Dächer können durch Brücken vernetzt werden. Durch treppenartige Anordnung von Gebäudeteilen können Verbindungen zur bodenständigen Grundflächen hergestellt werden.
Fassaden
Auch begrünte Fassaden gibt es schon lange, aber eher an alten Bauernhäuser auf dem Land als an mehrgeschossigen Stadthäusern, Bankhochhäusern und Industrieanlagen. Eine Möglichkeit: Flächenhafte Begrünungsmodule, die mit einfachen Mitteln an Fassaden angebracht werden können und die durch Anschluss an eine Bewässerungsanlage wartungsarm sind. Die Elemente können aus einem Gerüst bestehen, an dem mehrere auswechselbare Pflanzgefäße aufgehängt werden. Fensterfassaden könnten durch berankte Schnurgerüste – Hopfenfeldern vergleichbar – begrünt und beschattet werden.
Ein interessanter Vorschlag sind vorbegrünte Pflanzennetze. Solche „Urban Pergolas“ sollen als Verschattungssystem der Aufheizung von Fassaden entgegenwirken und die Städte in einen „diversen Großstadtdschungel“ verwandeln. Die Pflanzennetze können an einem oder zwischen mehreren Gebäuden angebracht werden und dadurch Grünflächen schaffen, ohne andere Nutzungen den Platz wegzunehmen (Urban Pergola 2021).
Balkone
Eine weitere Möglichkeit der vertikalen Begrünung, die in wenigen Beispielen schon verwirklicht ist, wäre die Ausgestaltung von Pflanzbalkonen mit Sträuchern und Bäumen (Boeri 2015).
Städte mit grünem Pelz
Ergänzend zu den genannten Maßnahmen können Verkehrswege, insbesondere Straßen und Schienenverkehr, wie U-Bahnen unter die Oberfläche verlegt werden, wodurch Platz für bodenständige Grünanlagen aber auch Rad- und Fußwege gewonnen würde. Regenwasser können den Zisternen gespeichert und in Trockenperioden zur Bewässerung genutzt werden wodurch die Kanalisation entlastet würde.
So könnten schließlich Städte entstehen, die ganz in einem grünen Pelz eingehüllt sind und die sich fast übergangslos in die umgebende Landschaft einfügen (vgl. Jean Nouvel 2014, Boeri 2015).
Begrünte Wohnblocks (Modellbau W.Probst)
Landwirtschaft
In der Landwirtschaft sollten großflächige Monokulturen durch ökologisch wertvollere Netze (Feldhecken, Blumenstreifen, Bachläufe) und Inseln (Feldgehölze, Feuchtgebiete) unterbrochen werden. Mischkulturen aus Gehölzen, mehrjährigen und einjährigen Nutzpflanzen (Agroforestry) könnten vor allem in wärmeren Klimaregionen eine ökologische Alternative zu Monokulturen darstellen. Die sehr aufwändige arbeitsintensive Bewirtschaftung würde durch einen Einsatz intelligenter Maschinen zu vertretbaren Produktionskosten möglich.
Nachhaltige Landwirtschaft: Vertical Farming spart Flächen und erleichter Stoffkreisläufe; Vernetzung durch Feldhecken und Wildpflanzenstreifen erhöht die Biodiversität in Agrarflächen und wird durch intelligente Maschinen möglich; Agroforestry, Anbau von Kulturpflanzen in mehreren Vegetationsschichten, fördert die Biodiversität und eignet sich vor allem für wärmere Klimazonen (z.B. in Kombination mit Kaffee- und Kakaoanbau) (Zeichung W.Probst)
Landwirtschaft 4.0
Lange Zeit wurden Landmaschinen – den Dinosaurier vergleichbar – immer größer und größer. Vergleicht man einen Traktor aus den 19hundertfünfziger Jahren mit einer heutigen Maschine wird dieser Hang zum Gigantismus deutlich. Er hängt natürlich direkt zusammen mit der Vergrößerung der landwirtschaftlichen Betriebee und vor allem der bewirtschafteten Flächen. Die Dinosaurier sind nicht zuletzt auch wegen ihrer Größe ausgestorben. Die immer größeren Landmaschinen stellen für die Landwirte eine große finanzielle Belastung dar und sicher sind sie ein Grund dafür, dass immer mehr landwirtschaftliche Betriebe aufgeben müssen. Auch die Verdichtung der Böden durch die Riesentraktoren ist ein großer Nachteil. Die Entwicklung kleiner intelligenter Landmaschinen könnte eine neue, ökologisch verträglichere und damit nachhaltigere Form der Landbewirtschaftung einleiten. Diese Maschinen könnten – ähnlich wie ein Schweizer Armeemesser – viele Funktionen in sich vereinen: ein Roboter, der jede Pflanze individuell behandelt, nicht nur mit Herbiziden, Insektiziden und Fungiziden, sondern auch mit angepassten Düngemitteln, und der auch für eine gezielte Bewässerung sorgt. Dies alles könnte in einem Arbeitsgang und in individuell angepassten Mengen geschehen. Die Folgen einer solchen Behandlung von Einzelpflanzen statt von ganzen Feldern bedeutet nicht nur eine deutliche Reduktion benötigter Chemikalien und anderer Ressourcen. Diese Maschinen könnten von Drohnen oder von Satelliten gesteuert die jeweiligen Zielorte erreichen. Eine Weiterentwicklung der Erntemaschinen könnte Mischkulturen und Agroforestry wirtschaftlicher machen.
Vertical Farming
Eine zukunftsweisende und flächensparende Form zur Produktion von Nahrungsmitteln und anderen nachwachsenden Rohstoffen wird mit dem Begriff „Vertical Farming“ bezeichnet. Der New Yorker Professor für Umweltgesundheit und Mikrobiologie Dickson Despommier entwickelte mit seinen Studenten ab 1999 entsprechende Ideen zunächst für die Nahrungsmittelversorgung der 50000 Einwohner Manhattans. Ausgangspunkt waren Überlegungen zum möglichen Gemüseanbau auf Dachflächen. In der Weiterentwicklung wurden Hochhäuser geplant, die insgesamt der Pflanzenkultur dienen sollen. Diese Einbindung von Farmen in das Innere von Gebäude wird mit dem Begriff „Sponge City- Architecture“ oder „Agritecture“ bezeichnet. In mehreren oder allen Stockwerken eines solchen Hochhauses sollen Pflanzen auf optimale Weise automatisch gesteuert und reguliert kultiviert werden. Gleichzeitig sind diese Kulturen in Kreislaufsysteme, insbesondere der Wasserwiederverwendung und Abwasseraufbereitung, eingebunden (Despommier 2011). Auch eine Kopplung mit Aquakulturen und anderen Formen der Nutztierhaltung ist möglich.
Der Vorteil solcher Plantscraper ist nicht nur der gegenüber normalem Farmland 10-20mal geringere Flächenverbrauch. Erhebliche Ressourcen könnten dadurch ein gespart werden, dass es einen geschlossenen Wasserkreislauf gibt und kontrollierte Umgebungsbedingungen den Einsatz von Pestiziden und Düngemitteln reduzieren. Die Kulturen sind unabhängig von Außenbedingungen wie Dürre, Frost, Starkniederschläge, Hagel und Sturm und sie können ganzjährig betrieben werden. Künstliches Licht kann Pflanzenwachstum rund um die Uhr auch in dunklen Jahreszeiten ermöglichen. Die schnellere und einfachere Versorgung der städtischen Bevölkerung mit frischen Nahrungsmitteln erfordert weniger Transportkosten, verbessert die Luft und mindert über Wasserspeicher die Überflutungsgefahr. Die Energieversorgung kann über Solarzellen, Windenergieanlagen und die Produktion von Biogas aus organischen Abfällen in einem Kreislaufsystem gesichert werden.
Der extrem dicht bevölkerte Stadtstaat Singapur plant seine Nahrungsmittelversorgung durch schwimmende Hochhäuser zu verbessern.
Voraussetzungen für den erfolgreichen Betrieb solcher Hochhausfarmen ist eine ausgefeilte Technik, die von intelligenten Computersystemen gesteuert wird. Das schwedische Architekturbüro Plantagon plant ein Forschungszentrum für urbane Landwirtschaft in Linköping zu entwickeln. Ausgangspunkt soll ein im Bau befindlicher Plantscraper sein, an dem technische Systeme erprobt und verbessert werden können.
Durch Brücken und Tunnel kann der Zerschneidungseffekt von Verkehrswegen gemindert werden (Zeichnung W.Probst)
Je dichter die Besiedelung, desto dichter sind nicht nur Städte, Siedlungen und Industrieanlagen, desto dichter ist auch das Netz von Verkehrswegen, insbesondere Straßen und Autobahnen (in Deutschland derzeit nach Erhebung des Umweltbundesamt knapp 20000 km², das entspricht rund 5,5% der Landesfläche). Das wirkt sich r nicht nur über den Flächenverbrauch und die Versiegelung sondern vor allem über den Zerschneidungseffekt nachteilig auf die Funktion von Ökosystemen aus. Mehr noch als Pflanzenarten sind Tierpopulationen durch die dadurch bedingte Verinselung betroffen. Auch die direkte Tötung von Tieren durch den Verkehr spielt eine Rolle. Indirekt wirkt sich dies über die Bestäuber und die Verbreitung von Früchten und Samen auf die Vegetation aus.
Eine Verbesserung kann einmal durch geeignetes Straßenbegleitgrün erreicht werden (Kühne/Freier 2012). Vor allem aber kann die trennende Wirkung von Verkehrsflächen durch Brücken, sowohl Brücken über schützenswerte Landschaftsteile als auch verbindende Grünbrücken, und Tunnel erreicht werden. Schutzgräben oder Zäune können in Kombination mit kleinen Tunneln insbesondere Amphibien bei ihren Laichwanderungen schützen (Krötenzaun, Krötentunnel). Nicht mehr benötigte Verkehrswege sollten renaturiert (entsiegelt) werden.
Schließlich sind die hohe Verkehrsdichte und die damit verbundenen Emissionen der Verkehrsmittel ein großes Problem. Sie wird einmal durch den Individualverkehr, zum anderen durch den Güterverkehr verursacht. Beide haben in den letzten Jahrzehnten ständig zugenommen. Eine größere Verlagerung dieses Verkehrs auf die Bahn wird schon lange als Ziel formuliert, ließ sich aber bisher politisch nicht durchsetzen. Auch eine Förderung dezentraler Produktion könnte der ständigen Zunahme des Güterverkehrs entgegenwirken.
Despommier, D. (2011): The vertical farm: Feeding the world in the 21th century. Picador (Nachdruck der Ausgabe von 2010)
De Souza Machado, A. A., Lau, C. W. u. a. (2019): Microplastics Can Change Soil Properties and Affect Plant Performance. In: Environmental Science & Technology. 53, S. 6044, doi:10.1021/acs.est.9b01339.
Dierkes, P., Homes, V. (2017): Artenschutz. UB 427 (41.Jg.), S. 2-11, Seelze: Friedrich
Kattmann, U. (1991). Bioplanet Erde: Neue Ansichten über das Leben. Unterricht Biologie, 15(162), 51-53.
Kattmann, U. (2004). Bioplanet Erde: Erdgeschichte ist Lebensgeschichte. Unterricht Biologie, 28(299), 4-14.
Keesing, F. u.a. (2010): Impacts of Biodiversity on the Emergence and Transmission of Infectious Diseases. Nature 468, S. 647-652
Kühne, S./Freier, B. (2012): Saumbiotope und ihre Bedeutung für Artenvielfalt und biologischen Pflanzenschutz. Workshop „Biological Diversity in Agricultural Landscapes“ – February 09-10, 2012, Berlin-Dahlem
Le Roy, L. G. (1973): Natur ausschalten – Natur einschalten. Stuttgart: Klett Cotta
Little, A. (2019): The fate of food. What we’ll eat in a bigger, hotter, smarter World. London: Oneworld Publications
Liu, Xiaoping et al. (2020): High spatiotemporal resolution mapping of global urban change from 1985 to 2015: Nature Sustainability 3, pp.564-570.
Mantyka-Pringle, C. S., Martin, T. G., Rhodes, J. R. (2012): Interactions between climate and habitat loss effects on biodiversity: a systematic review and meta-analysis. Global Change Biology 18, pp. 1239-1252
Shah, S. (2020): Woher kommt das Corona-Virus? Le Monde diplomatique vom 12.03.2020
Smil, V. (2019): Growth – From microorganismes to megacities. Cambridge MA.: MIT-Press
Trommer, G. (1994): Didaktisch differenzierte Leitbilder – ein Drei-Umwelten-Modell zum pägagogischen Umgang mit Natur und Landschaft. Workshop Ökologische Leitbilder, Cottbus 9.6.1994. TUC Aktuelle Reihe 6/94:57-62
Das Berliner Stadtschloss – korrekt Königliches Schloss – steht nun wieder, zumindest seine Fassade. Innerlich hat es sich erneuert. Das Humboldt-Forum soll Weltoffenheit repräsentieren: „Neben dem Ethnologischen Museum Berlin und dem Museum für Asiatische Kunst der Staatlichen Museen wird es auch die Berlin-Ausstellung des Stadtmuseums und das Humboldt-Labor der Humboldt-Universität beheimaten“ (Wikipedia). Doch Schlossbau und Schlossinhalt sind nach wie vor sehr umstritten. Die Wellen in den Kulturspalten der Zeitungen und in den Kommentaren der Medien und sozialen Netze schlagen hoch.
Bevor das Schloss nach dem Muster des alten Preußenschlosses wieder aufgebaut wurde und nachdem der Palast der Republik abgerissen war, habe ich hier auf meiner Homepage einen Vorschlag für die Gestaltung dieses Freiraumes gemacht. Aus aktuellem Anlass stelle ich ihn noch einmal ein, weil ich nach wie vor meine, so eine „Pyramide für Berlin“ wäre eine schöne Alternative gewesen. Nun ist es natürlich zu spät, aber in der Zukunft wird es um ähnliche Entscheidungen gehen. Ich hoffe, man wird sich dann erinnern und zukunftsweisendere und nachhaltigere Lösungen suchen, anstatt alte Fassaden wieder zuerrichten.
Eine Pyramide für Berlin
Das Stadtschloss von Berlin stand mit seinen ältesten Teilen etwa 500 Jahre lang, der Palast der Republik gerade 30 Jahre. An dem Platz, an dem diese Monumentalbauten das Zentrum eines jeweils nicht unumstrittenen Staatswesens markierten, dehnt sich heute eine Rasenfläche. Mitten im betriebigen Zentrum Berlins lädt sie zum Picknick machen und Ausruhen ein. Nachdem das Stadtschloss mit seiner 60 m hohen Kuppel im Zweiten Weltkrieg zerbombt und von den DDR- Sozialisten später vollends abgerissen wurde, und nach dem auch der Palast der Republik aus ideologischen und bautechnischen Gründen (Asbest!) der Abrissbirne zum Opfer fiel, soll das Schloss in seiner alten äußeren Form wieder aufgebaut werden.
Die Wiese am 16. Juni 2010
Zunächst frage ich mich natürlich: Warum nicht viel Geld sparen und diese gegenwärtige, bürgerfreundliche Situation auf sich beruhen lassen, überdauern lassen? Einen Rasen kann man nicht abreißen und auch mit Bomben nur schwer zerstören, man kann ihn höchstens wachsen lassen. Wenn ihn niemand mehr mäht und niemand mehr abweidet, kann es sein, dass mit der Zeit ein Wald aus ihm wird. Dann könnte man immerhin wieder mit einer Motorsäge größere Veränderungen bewirken. Aber die Entscheidung darüber würde frühestens in einigen Jahrzehnten nötig.
Das wäre zu einfach, meinen Sie? Berlins Mitte erfordere eine aufwändigere architektonische Lösung, es müsse ein neues Zentrum her, ein Ziel für die Prachtstraße „Unter den Linden“, ein markanter Ort, ein Wahrzeichen für die Stadt? Nun – da würde ich den Bau einer Pyramide vorschlagen. Die ältesten Monumentalbauten der Menschheit, die Pyramiden von Gizeh, stehen heute, etwa 6500 Jahre nach ihrem Bau, immer noch – immerhin schon 13 mal so lang wie das Schloss und über 200 mal so lang wie der Palast. Ich stelle mir allerdings keine Steinpyramide vor, sondern eine Graspyramide. Man könnte sie allmählich wachsen lassen mit dem Aushub von Baugruben, U-Bahn-Bauten, unterirdischen Bahnhöfen, Abwasserleitungen und anderweitig anfallendem Aushub- und Abraummaterial. Mit der Pyramidenform würde man einer uralten menschlichen Bau- und Konstruktionsidee folgen, der grüne Bewuchs wäre Symbol für die Symbiose von Kultur und Natur. Je nach zur Verfügung stehenden Mitteln wäre es überdies ein leichtes, Gebäude und andere Konstruktionen in die Pyramide zu integrieren. Außer einem Spazierweg zur Pyramidenspitze könnte man sich allerhand andere Einrichtungen vorstellen, wie Rutschbahnen, Seilbahnen, Mountainbikestrecken und Ähnliches. In der Mitte der Pyramide würde, durch einen Tunnel erreichbar, ein Aufzug für Rollstuhlfahrer und Gehbehinderte eingebaut.
Die grüne Pyramide würde sich allmählich verändern, Büsche würden Wachsen, später auch Bäume …
Eine Fahne auf der Pyramidenspitze könnte verschiedenen Zwecken dienen, zum Beispiel könnte man damit anzeigen, dass der Bundespräsident in der Stadt ist. Durch entsprechende Nationalitätenflaggen könnte man über Staatsbesuche informieren, aber auch Feste und Gedenktage könnten durch fantasievolle Beflaggung signalisiert werden.
Wäre das nicht schöner als ein nachgemachter Prachtbau, der vielleicht noch ins 19. Jahrhundert passte? Schade, die Entscheidung ist wohl gefallen – einzige Chance für die Idee: das Geld geht aus!
Die Tourist-Information und der Arbeitskreis Tourismus laden Feriengäste und Einheimische ein:
Botanischer Spaziergang
mit Prof. Dr. Wilfried Probst
Samstag, 12.09.2020, 10:00 Uhr
Treffpunkt: Wanderparkplatz bei Appenweiler an der Straße zwischen Appenweiler und Brochenzell, links der Straße etwa 300 m hinter Appenweiler.
Ziel ist das Waldgebiet am westlichen Abhang des Schussensbeckens, an dieser Stelle als Weißenauer Wald bezeichnet. Es zeichnet sich durch einen sehr abwechslungsreichen Baumbestand aus. Auf unserer kleinen Runde werden wir uns besonders mit den Pflanzen an den feuchten Wegrändern und Gräben befassen. Vielleicht finden wir auch einige Pilze.
Bitte beachten Sie die bekannten Hygiene- und Schutzmaßnahmen sowie den Mindestabstand von 1,5 m. Die Teilnehmer sind verpflichtet, ihre Kontaktdaten zu hinterlassen.
Dauer ca. 2,5 Stunden. Die Veranstaltung ist kostenlos.
Teilnahme auf eigene Gefahr.
Weg von Oberteuringen zum Exkursionsgebiet. Ausschnitt aus den Topographischen Karten 1: 25.000 Markdorf 8222 und Ravensburg 8223Appenweiler mit Treffpunkt im Luftbild (Google Earth)
Wälder am Rand des Schussenbeckens
Größere zusammenhängende Waldgebiete sind im heutigen Oberschwaben ziemlich selten. Im Laufe der mehrtausendjährigen Siedlungsgeschichte ist die typische kleinräumige Landschaft aus Feldern – heute vielfach Obstplantagen -, Wiesen und kleinen Waldstücken entstanden. Das größte Waldgebiet, der Altdorfer Wald etwa zwischen Vogt und Wolpertswende gelegen, hat immerhin eine Längsausdehnung von ca. 17 km. Dagegen ist das Waldgebiet , das sich am westlichen Rand des Schussenbeckens etwa von Ravensburg bis Meckenbeuren erstreckt, mit knapp 8 km deutlich kleiner. Die geplante Umgehungsstraße für Meckenbeuren könnte es noch weiter verkleinern. Trotzdem kann man in diesem Wald stundenlang wandern. Mehrere Bäche entwässern das Gebiet zur Schussen hin. Sie haben sich zum Teil ziemlich tief in die Jungmöräne eingeschnitten.
Die Artenzusammensetzung des Baumbestandes ist recht abwechslungsreich. Neben Buchen, Eichen und anderen Laubbäumen finden sich auch Tannen, Fichten und Kiefern, bachbegleitend Eschen und Erlen. Typische Waldgesellschaft ist ein Buchen-Tannen-Wimpernseggenwald. Aber der Untergrund ist recht abwechslungsreich und dies wirkt sich auch auf die Vegetation aus.
Mit dem Bodenseebecken wird das untere Schussental im Allgemeinen zum Landschaftsraum Bodensee-Schussen-Becken zusammengefasst, der am Ende der Würmeiszeit durch den Rheingletscher geprägt wurde. Das aus Satellitenaufnahmen errechnete digitale Geländemodell zeigt sehr gut die Ausdehnung der Vereisung am Ende der letzten Eiszeit. KS markiert den Gletscherrand zur Zeit des sogenannten Konstanzstadiums vor etwa 15.000 Jahren.
Vereisungsgebiet des Rheingletschers im Bereich des Bodensees. Schwarze Linie: Schussen; weiß punktiert (IWEM): Innere Würmendmoräne; weiß gestrichelt (KS): Gletscherrand des Konstanzer Stadiums (vor ca. 15.000 J); dünne weiße Linien: Moränenrelikte innerhalb der Endmoräne des Konstanzer Stadiums; Beschriftung von West nach Ost: KON Konstanz; ÜBE Überlingen; SAL Salem; MAR Markdorf; GEH Gehrenberg; FRI Friedrichshafen; TET Tettnang; RAV Ravensburg. Quelle: Beckenbach, E., Müller, T., Seyfried, H., Simon, T. (2014): Potential of a high-resolution DTM with a large spatial coverage for visualization, identification and interpretation of young (Würmian) glacial morphology. Quarterny Science Journal 63 (2),pp. 107-129. Zahlreiche ähnliche Abbildungen finden sich in Seyfried, H., Simon, T., Beckenbach, E., Müller, T. (2019): Der Südwesten im digitalen Geländemodell. Wie LitDAR-Daten unsere Sicht auf die Welt verändern. Sonderbände der Gesellschaft für Naturkunde in Württemberg e.V. Bd. 4, Verlagsdruckerei Schmidt, 914 13 Neustadt an der Aisch
Das Waldgebiet ist von einem relativ dichten Wegenetz durchzogen. . Diese Wege dienen vor allem der forstlichen Bearbeitung. Oft sind sie von Entwässerungsgräben begleitet, denn in der Jugendmoräne sind immer wieder Lehmschichten eingelagert, die zu Staunässe und Quellhorizonten führen. Deshalb ist die wegbegleitende Vegetation recht vielseitig. Besonders auffällig ist das an vielen Stellen sehr feuchtigkeitsbedürftige Pflanzen gedeihen.
Der Spaziergang
Exkursionsweg. Ausschnitt aus der Topographischen Karte 1: 25.000 Ravensburg 8223
Wir beschäftigen uns zunächst mit dem hochgewachsenen Maisfeld gegenüber dem Wanderparkplatz am Waldrand. Die Kulturen dieses sehr wuchskräftigen Grases haben im letzten Jahrzehnt sehr stark zugenommen. Die Pflanzen werden vor allem zur Herstellung von Silagefutter und zur Biogasproduktion genutzt. Sie erfordern hohen Dünger-und Pestizideinsatz. Vor allem vor dem Auflaufen im Frühjahr (Anfang Mai) werden die Unkräuter mit Herbiziden zurückgehalten. Der Rand dieses Maisfeldes ist aber trotzdem sehr unkrautreich.
Wir beobachten drei sehr charakteristische Gräser:
Grüne Borstenhirse (Setaria viridis),
Hühnerhirse (Eragrostis crus-galli) und
Blut-Fingerhirse (Digitaria sanguinalis
Borstenhirse, Hühnerhirse und Fingerhirse im Maisfeld (Foto W. Probst,13.9.2020)
Weitere Arten:
Ampfer-Knöterich (Persicaria lapathifolia)
Weißer Gänsefuß (Chenopodium album)
Stumpfblättiger Ampfer (Rumex obtusifolius)
Kohl-Gänsedistel (Sonchus oleraceus)
Kleinblütiges Knopfkraut bzw. Franzosenkraut (Galinsoga parviflora)
Gänse-Fingerkraut (Potentilla anserina)
Löwenzahn (Taraxacum officinale agg.)
Echte Zaunwinde (Calystegia sepium)
Spitz-Wegerich (Plantago lanceolata)
Breitblättriger Wegerich (Plantago major)
Große Brennnessel (Urtica dioica)
Wiesen-Bärenklau (Heracleum sphondylium)
Steifer Sauerklee (Oxalis stricta)
Acker-Minze (Mentha arvensis)
Als Besonderheit wächst hier an mehreren Stellen die aus China stammende Quirl- oder Gemüse-Malve (Malva verticillata), deren Blätter in China seit mehr als 2500 Jahren als Gemüse genutzt werden. Die Pflanze findet sich seit einiger Zeit in den Blühstreifen an Ackerrändern, da sie teilweise Bestandteil der dafür vorgesehenen Saatmischungen ist.
Quirl- oder Gemüse-Malve (Malva verticillata) an Maisfeld bei Appenweiler (Fotos S. und W.Probst, 13.9.2020)
Nun folgen wir in langsamem Botanikertempo dem Weg in den Wald hinein. Alle Teilnehmer* innen erhalten Kärtchen mit Namen von Baum-und Straucharten, die in diesem Waldteil zu finden sind. Gefunden werden:
Gewöhnliche Esche (Fraxinus excelsior) – Blätter und Seitenzweige gegenständig –
Vogelbeere, Eberesche (Sorbus aucuparia) – Blätter und Seitenzweige wechselständig –
Stiel-Eiche (Quercus robur) – Früchte lang gestielt, die der Trauben-Eiche fast sitzend –
Rot-Buche (Fagus sylvatica) – glatte Rinde, keine Borkenbildung –
Schwarz-Erle (Alnus nigra) – Blätter stumpf, ohne Blattspitze –
Berg-Ahorn (Acer pseudoplatanus)
Winter-Linde (Tilia platyphyllos) – im Gegensatz zur Sommer-Linde mit bräunlichen Härchen in den Winkeln der Blattadern auf der Blattunterseite –
Wald-Kiefer (Pinus sylvestris)
Rot-Fichte (Picea abies)
Weiß-Tanne (Abies alba)
Sträucher:
Weißdorn (Crataegus spec.) – es gibt mehrere sehr ähnliche Arten –
Gewöhnliche Hasel, Haselstrauch (Corylus avellana) – die Kätzchen für das nächste Jahr sind schon angelegt –
Schlehe, Schwarzdorn (Prunus spinosa) – viele blaue Steinfrüchte, die Seitenzweige laufen fast in einem rechten Winkel ab, dadurch ist Schlehengebüsch besonders undurchdringlich –
Schwarzer Hollunder (Sambucus nigra)
Echter Schneeball (Viburnum ebulus) – die roten Beeren sind leicht giftig –
Blutroter Hartriegel (Cornus sanguinea) – „blutrot“ bezieht sich auf die Herbstfärbung –
Faulbaum, Pulverholz (Rhamnus frangula) – der Name bezieht sich auf den leichten Fäulnisgeruch der Rinde, die als starkes Abführmittel gilt; die hochwertige Holzkohle wurde früher bevorzugt zur Schwarzpulverherstellung verwendet; Futterpflanze für die Raupen des Zitronenfalters –
Unterschiede von Fichte und Tanne (aus: Probst, W. (2007): Pflanzen stellen sich vor. Köln: Aulis)
An einer Wegkreuzung steht eine hohe Weiß-Tanne, die im oberen Kronenbereich zahlreiche Tannen-Misteln trägt. Diese Mistel ist eine Unterart (Viscum album subsp. abietis), die nur an Tannen vorkommt. Misteln sind Halbschmarotzer, die ihrem Wirt Wasser und Mineralstoffe entziehen, aber selbst über Photosynthese Nährstoffe produzieren. Ihre weißlichen Beerenfrüchte enthalten einen sehr klebrigen Schleiml, aus dem man früher Vogelheim hergestellt hat. Sie werden gerne von Drosseln gefressen (daher der Name der Mistel-Drossel) „Turdus ipse sibi cacat mortem“ –„Die Drossel scheißt sich selbst den Tod“ (römisches Sprichwort).
Neben der Tanne steht eine Rot-Fichte. Unterschiede zwischen Fichte und Tanne werden besprochen.
Weiß-Tanne (Foto W. Probst,13.9.2020)
Rot-Fichte (Foto W- Probst 13.9.2020)
Wir folgen dem Weg bis zur Kreuzung mit einem Betonsträßchen, auf das wir dann nach links abbiegen. Der Weg ist von einem mehr oder weniger tiefen Graben begleitet, deshalb finden sich in der wegbegleitenden Vegetation häufig besonders feuchtigkeitsliebenden Pflanzen:
Pfeifengras, Benthalm (Molinia caerulea) – die langen knotenlosen Stiele des Blütenstandes wurden früher zum Pfeifen reinigen verwendet; typisches Gras der Pfeifengraswiesen –
Steife Segge (Carex elata)
Zittergras-Segge (Carex brizoides)
Hänge-Segge (Carex pendula)
Flatter-Binse (Juncus effusus) – das Tragblatt des Blütenstandes sieht wie eine Fortsetzung der Sprossachse aus –
Gilbweiderich (Lysimachia vulgaris) – Knoten meistens mit drei Blättern, seltener auch zwei oder vier –
Blutweiderich (Lythrum salicaria) – nicht näher mit dem Gilbweiderich verwandt, „-weiderich“ bezieht sich auf die weidenartigen Blätter beider Pflanzenarten –
Echtes Mädesüß (Filipendula ulmaria) – die süßlich duftenden Blüten wurden früher zur Aromatisierung von Bier und Wein verwendet, Name eventuell von „ Met-Süße“; die ganze Pflanze aber besonders der Wurzelstock enthält Methylsalicylat, das wegen seiner desinfizierenden Wirkung zum Beispiel Zahnpasta und Kaugummis beigefügt wird; der Name „Aspirin“ leitet sich von der früheren Bezeichnung „Spiraea ulmaria“ und dem A von Acetylsalicylsäure ab.
Gewöhnliches Hexenkraut (Circaea lutetiana)
Lippenblütler:
Wasser-Minze (Mentha aquatica) – eine der drei Stammarten der Pfeffer-Minze (Mentha x piperita, x für Hybridart)
Ross- Minze (Mentha longifolia) – weitere Stammart der Pfefferminze –
Ufer-Wolfstrapp (Lycopus europaeus) – die tief gesägten, gegenständigen Blätter erinnern an ein Wolfseisen –
Wald-Ziest (Stachys sylvatica) – starker, etwas unangenehmer Geruch –
Wiesen-Bärenklau (Heracleum sphondylium) – nicht zu verwechseln mit dem wesentlich größeren, aus dem Himalaja eingeschleppten Riesen-Bärenklau, der wegen des Kontaktgiftes in seinen Blättern gefürchtet wird. Das Gift wirkt vor allem zusammen mit UV-Licht. „Ist der Stängel kantig rau, heißt die Pflanze Bärenklau“ –
Wald-Engelwurz (Angelica sylvestris) – mehrfach zusammengesetzte, sehr große Blätter und auffällig große Blattscheiden –
Wald-Sanikel (Sanicula europaea)
Gewöhnliche Bibernelle (Pimpinella saxifraga) – zur gleichen Gattung gehört Anis (Pimpinella anisum) aus dem Balkan
nur Blätter sehen wir vom
Wiesen-Kerbel (Anthriscus sylvestris),
Giersch (Aegopodium podagraria) und
Behaarter Kälberkropf (Chaerophyllum hirsutum)
Da nun schon mehr als die Hälfte der Zeit um ist, die wir uns für unseren Spaziergang vorgenommen haben legen wir den Weg auf dem Betonsträßchen etwas schneller zurück. Besondere Beachtung schenken wir einem wassergefüllten Graben mit fruchtendem Breitblätterigen Rohrkolben (Typha latifolia). Das Betonsträßchen endet an einer Wegkreuzung, in deren Nachbarschaft sich ein pilzförmiger Unterstand befindet. Im grasigen Wegrand blühen einige Herbst-Zeitlosen (Colchicum autumnale) . Diese in allen Teilen hochgiftige Art ist die einzige ihrer Gattung in Mitteleuropa. Verbreitungsschwerpunkt der Gattung sind die Trocken- und Halbtrockengebiete Vorder- und Westasiens, an deren Klimabedingungen die Arten mit ihrer unterirdischen Speicherknolle besonders gut angepasst sind. Das enthaltene Gift Colchicin verhindert die Ausbildung korrekter Mitosespindeln und damit eine geordnete Zellteilung.
Unser weiterer Spaziergang folgt nun nach links einem kleineren Waldweg.
Wir besprechen, wie man mit einem Zollstock, einem Bindfaden und eventuell noch einem Maßband das Holzvolumen eines Baumes und damit auch den von ihm gespeicherten Kohlenstoff abschätzen kann. Die einzelnen stehenden Stämme von Kiefern und Tannen auf einer großen Lichtung, die vermutlich durch Borkenkäfer verursachten Kahlschlag entstanden ist, bieten sich zwar für diese Untersuchung an. Angesichts der fortgeschrittenen Zeit entschließen wir uns aber, auf die praktische Ausführung zu verzichten.
Abschätzung der in einem Baum gespeicherten Kohlenstoffmenge
Entlang des Weges zieht sich eine feuchte Niederung mit dichtem Seggenbestand (vermutlich die lange Ausläufer bildende Schlank-Segge (Carex acuta). Außerdem beachten wir die großen, spIrrigen Blütenstände der Wald-Simse (Scirpus silvaticus).
Leider haben wir auf unserem Spaziergang keine Pilze gefunden. Aber als ich einen Tag später mit meiner Frau noch einmal in dem Gebiet war, sahen wir Graue Lärchenröhrlinge (Suillus viscidus), brauchbare Speisepilze.
Scan des Grauen Lärchenröhrlings (Suillus viscidus) ; Hutdurchmesser 5,5 cm (W. Probst, 13.9.2020)
Die Tourist-Information und der Arbeitskreis Tourismus laden Feriengäste und Einheimische ein:
Botanischer Spaziergang
mit Prof. Dr. Wilfried Probst
Samstag, 08.08.2020, 10:00 Uhr
Ersatztermin bei schlechtem Wetter: 15.08.2020
Treffpunkt: Wanderparkplatz westlich von Raderach am Drumlin Heidengestäud.
Kopie aus den Topografischen Karten 1:25 000 Markdorf 8222 und Friedrichshafen 8322 des Landesvermessungsamtes Baden-Württemberg
Unser Spaziergang führt uns in die Raderacher Drumlinlandschaft. Rund um den Drumlin „Heidengestäud“, auf dessen Gipfel sich ein möglicherweise prähistorischer Ringwall befindet, bietet der Weg einen sehr schönen Ausblick auf das Hepbach-Leimbacher Ried. Auf unserer kleinen Runde achten wir besonders auf die sehr artenreiche Flora der Wegränder. Wir werden aber auch eine neue Kulturpflanze kennenlernen.
Für empfindliche Personen empfiehlt sich die Mitnahme eines Mückenschutzmittels.
Bitte beachten Sie die bekannten Hygiene- und Schutzmaßnahmen sowie den Mindestabstand von 1,5 m. Die Teilnehmer sind verpflichtet, ihre Kontaktdaten zu hinterlassen.
Dauer jeweils ca. 2,5 Stunden. Die Veranstaltung ist kostenlos.
Teilnahme auf eigene Gefahr.
Drumlins
Raderacher Drumlinlandschaft, 22.6.2013 (Foto W. Probst)
Die Landschaft um Raderach ist gekennzeichnet durch viele langgestreckte Hügel, die im Profil etwa wie ein umgekehrter Löffel aussehen, mit einer etwas steileren und einer länger ausgezogenen Seite. Diese in der letzten Eiszeit entstandenen Formen werden mit einem irischen Ausdruck als Drumlin bezeichnet (von irisch „Druim“ „schmaler Rücken“). Ihre Ausrichtung entspricht der Fließrichtung des Gletschers (Abbildung). Über ihre Entstehung gibt es unterschiedliche Theorien. Wenn man annimmt, dass der Gletscher über ein plastisches Grundmoränenmaterial aus alten Sedimenten fließt, kann man sich gut vorstellen, dass kleine Erhebungen im Untergrund zu einem Aufstau führen. Größe und Form hängen dann vom Material, von der Fließrichtung des Gletschers und von der Geschwindigkeit des Abtauens ab. Häufiges auftreten von Drumlinfeldern – wie in der Umgebung von Raderach – gilt als Indiz für ein rasches Abtauen.
Entstehung eines Drumlins Im Google-Earth-Bild kann man das Raderacher Drumlinfeld sehr gut erkennen, da die Hügel im Gegensatz zum Umland bewaldet sind
Heidengestäud
Der kleine Berg, um den uns unser Spaziergang herumführen wird, ist in der topographischen Karte 1: 25.000 TK 8222 Markdorf als „Heidengestäud“ bezeichnet. In der Liste von Burgen und Schlössern in Baden-Württemberg wird die Wallanlage unter Heidengestied genannt. Dass solche Bergkuppen und speziell auch Drumlins für Befestigungsanlagen genutzt wurden, ist nicht ungewöhnlich. Ob es sich bei diesem Rest einer Wallanlage tatsächlich um eine alte keltische Viereckschanze handelt, ist nicht unumstritten. In diesem Fall wäre sie mindestens 2500 Jahre alt. Möglicherweise wurde das Bauwerk aber auch erst später im Mittelalter errichtet.
Auch der Ort Raderach wurde auf einem Drumlin erbaut, und zwar an einer Stelle, an der im 13. Jahrhundert eine Burg errichtet worden war. Die Steine der Burg wurden zum Teil für den Bau des heutigen Gasthofs Krone verwendet.
Der Spaziergang
Der Exkursionsweg (Auschnitt aus TK 8222 Markdorf)
Wir folgen zunächst dem Weg am Waldrand entlang. Die Wiese rechts vom Weg wurde leider gerade gemäht, der zweite Schnitt in diesem Jahr. Von den Wiesenpflanzen sieht man deswegen nur noch wenig an den Wegrändern. Die auffälligste Wiesenpflanzen, die kurz vor dem Schnitt den Aspekt der Wiese bestimmt hat, ist aber auch am Wegrand reichlich vorhanden: Wiesen-Bärenklau (Heracleum sphondylium). Sie kommt vor dem ersten Schnitt normalerweise nicht zum Blühen, treibt dann aber danach noch einmal aus und wird vor dem zweiten Schnitt die höchste Pflanze der Wiese. Empfindliche Leute können mit einer leichten Hautallergie auf die Berührung mit Wiesen-Bärenklau reagieren (Wiesen-Dermatitis), kein Vergleich allerdings mit den heftigen Reaktionen auf den Riesen-Bärenklau (Heracleum mantegazzianum).
Weitere Arten von Wiesen- und Wegrand: Scharfer Hahnenfuß, Gewöhnliche Schafgarbe, Weiße Taubnessel, Gefleckte Taubnessel, Wirbeldost, Gewöhnliches Johanniskraut, Nesselblättrige Glockenblume, Gewöhnlicher Rainkohl, Kleiner Pippau, Wolliges Honiggras.
Die Durchwahsenblättrige Silphie ist eine neue Nutzpflanze in Mitteleuropa, die vor allem der Biomasseproduktion dient
Bei 1 kommen wir an ein Feld mit sehr hohen, gelb blühenden Pflanzen. Die Blüten sehen ein bisschen wie kleine Sonnenblumen aus. Es handelt sich um die Durchwachsenblättrige Silphie (Silphium perfoliatum), ein aus Nordamerika stammender Korbblütler. Die Pflanze wird seit einigen Jahren als ergiebiger Biomasseproduzent in Mitteleuropa angebaut. Sie lässt sich auch als Futter- und Silagepflanze und als Bienenweide nutzen.
Das besondere Merkmal der bis 3 m hoch wachsenden, mehrjährigen Pflanze sind die gegenständigen, am Stängel zu Paaren verwachsenen Blätter, die einen Becher bilden. Der im englischen gebräuchliche Name „cup plant“ wurde deshalb auch ins Deutsche übernommen: Becherpflanze. (Foto J.Probst, 8.8.2020)
Die Silphie kann zehn Jahre lang am gleichen Standort geerntet werden und produziert ab dem zweiten Jahr mit 13-20 t Trockenmasse pro Hektar sogar eher mehr als Mais (http://bizz-energy.com/biogasbranche_wirbt_f%C3%BCr_us_pflanze) . Dass sie sich gegenüber Mais bisher trotzdem noch nicht durchgesetzt hat liegt vor allem daran, dass die Kosten im ersten Anbaujahr verhältnismäßig hoch sind. Im ersten Jahr bilden sich nur grundständige Rosetten, die schnell von Unkraut überwuchert werden und dieses Unkraut kann weitgehend nur mechanisch bekämpft werden. Ab dem zweiten Jahr ist jedoch die Beschattung des Bodens so stark, dass Unkräuter zurückgehalten werden. Eine kostengünstige Lösung wäre es deshalb, im ersten Jahr eine Mischkultur mit Mais anzulegen. Dadurch, dass die Kulturen über mehrere Jahre bestehen bleiben können, ist auch die Bodenerosion gering.
Zwei Stammarten von Kulturpflanzen
Kompass-Lattich (Lactuca serriola) (Foto W. Probst, 2016)
Der Weg geht nun steiler bergauf, am Hang zum kleinen Drumlin rechts ist ein großes Silphienfeld zu sehen. Links am Wegrand fällt uns eine Pflanze mit fast senkrecht stehenden, tief gebuchten und etwas bestachelten Blättern auf, die eine reich verzweigten rispigen Blütenstand mit vielen kleinen Korbblüten entwickelt hat. Der Kompass-Lattich oder Stachel-Lattich (Lactuca serriola) hat seinen Namen daher, dass er an sonnigen Standorten seine Blätter senkrecht und weitgehend in Nord-Süd-Richtung ausrichtet, wodurch sie vor der starken Sonneneinstrahlung geschützt werden. Er gilt als die Stammpflanze des Grünen Salates (Lactuca sativa). Die genetische Ähnlichkeit der beiden Arten ist so groß, dass man heute davon ausgeht, dass eine Aufspaltung in zwei Arten nicht gerechtfertigt ist.
Die zweite Art, ebenfalls ein Korbblütler, hat schöne hellblaue Blütenköpfe in lockeren Blütenständen. Der deutsche Name „Wegwarte“ (Cichorium intybus) bezeichnet ihren typischen Standort an Wegrändern. Die Wildpflanze, nach ihrem wissenschaftlichen Namen auch „Zichorie“ genannt, wird bis heute in einigen Gegenden des Mittelmeergebietes als Salat oder Gemüse verwendet. Sie ist auch eine traditionelle Heilpflanze zur Appetitanregung und Stimulierung von Verdauungssäften. Als Wurzelzichorie hat man aus ihren gerösteten Pfahlwurzeln Kaffeeersatz hergestellt (Zichorien-Kaffee). Erst im 19. Jahrhundert wurde aus Zichorien der Chicoréesalat gezüchtet : „Nach einer Überlieferung zog der Chefgartenbauer am Botanischen Garten in Brüssel, Bresier, 1846 die ersten Chicoréesprossen. Die Wurzeln ließ er zwar noch im Freiland wachsen, zum Sprossen verhüllte er sie jedoch lichtdicht, so dass sie möglichst wenig Bitterstoffe entwickelten. Nach einer anderen Version soll diese Art des Treibens auf eine zufällige Beobachtung zurückgehen: Als belgische Bauern 1870 ihre Zichorienwurzeln infolge ungewöhnlich hoher Ernte im Gewächshaus einschlugen, entdeckten sie während des Winters die kräftigen Knospen.“ (Wikipedia, 12.8.2020).
Auch der Endiviensalat gehört zur Gattung der Wegwarten oder Zichorien. Die Heimat von Cichorium endivia ist das Mittelmeergebiet. Heute werden weltweit zahlreiche Endiviensalatsorten angebaut.
Bei 2 erreichen wir einen schönen Aussichtspunkt mit Blick auf den Gehrenberg, die Orte Hepbach und Leimbach und das davor liegende Naturschutzgebiet Hepbach-Leimbacher Ried, das von einer Heckrinder-Herde beweidet wird. Bei dem mit gut 750 m ü.NN hohen Gehrenberg handelt es sich um keinen Drumlin. Er ist durch Erosion während der letzten Vereisung entstanden. Nach der vorletzten Vereisung haben sich in tiefer gelegenen Gebieten Schotter abgelagert, die durch kalkhaltiges Wasser verfestigt wurden. Dieser harte eiszeitliche Nagelfluh bot bei der letzten Vereisung einen erhöhten Widerstand gegen Erosion und so kam es zur Reliefumkehr: frühere Täler wurden zu Bergen.
Blick aufs Hepbach-Leimbacher Ried am 27.4.2012 (Foto W. Probst)
Die Panoramakarte zeigt ein Landschaftsbild, bei dem sich der Gletscher etwa auf eine Linie vom Schussenbecken bei Ravensburg bis Markdorf zurückgezogen hat. Dabei hat sich im Bereich des heutigen Hepbacher-Leimbacher Rieds ein Eisstausee gebildet, aus dem sich das Ried mit seinen bis 10 m mächtigen Torfschichten entwickelt hat. Heute liegt dort die Quelle der Brunnisach.
Autor: Thommi Gitter, entnommen aus: Markdorf, Geschichte und Gegenwart, 1990Wegränder mit Hochstauden
Wir gehen dann ein kleines Stück zurück und folgen dem Weg in den Wald hinein, der hier von relativ jungen Bäumen mit einem hohen Anteil an Berg-Ahorn gebildet wird. Der Weg wird gesäumt von einer breiten Hochstaudenflur 3. Bemerkenswerte, besonders große Stauden sind der giftige Pferde-Hollunder (Sambucus ebulus), im Gegensatz zu seinen Schwesterarten Schwarzer Hollunder und Trauben-Hollunder kein Gehölz sondern eine krautige Pflanze; Wald-Engelwuz (Angelica sylvestris), Behaarte Karde (Dipsacus pilosus), außerdem Acker-Kratzdistel (Cirsium arvense), Lanzett-Kratzdistel (Cirsium vulgare), Kohl-Kratzdistel (Cirsium oleraceum), Indisches Springkraut (Impatiens glandulifera), Wiesen-Bäremnklau (Heracleum sphondylium), Große Brennnessel (Urtica dioica), Nesselblättrige Glockenblume (Campanula trachelium) und Zottiges Weidenröschen (Epilobium hirsutum).
Auf der linken Seite es Weges ist die Vegetation teilweise weniger üppig. An dem Grabenrand entdecken wir Echtes Tausendgüldenkraut (Centaurium erythrophyllum).
Weitere Arten sind Taumel-Kälberkropf (Chaerophyllum temulum), Gewöhnlicher Dost oder Oregano (Origanum vulgare), Kanadische Goldrute (Solidago canadensis). Die Rasen-Schmiele (Deschampsia cespitosa) deutet auf einen Quellhorizont hin. Auch einzelne Bulte des Pfeifengrases (Molinia caerulea) und Wasserdost (Eupatorium cannabinum) zeigen feuchten Untergrund an.
Kaisermantel auf dem Blütenstand einer Behaarten Karde (Foto J. Probst, 8.8.2020)
Der Weg führt nun wieder bergab in einen schönen Buchen-Hochwald (Waldmeister-Buchenwald). Die übrigen Hochstauden erhalten hier nicht mehr genügend Licht aber einen Bestand von Hexenkraut (Circaea lutetiana) und Kleinblütigem Springkraut (Impatiens parviflora) reicht das Licht aus.
Fliege (cf. Mesembrina maridiana) auf Kanadischer Goldrute (Solidago canadensis) Foto: J. Probst, 8..8.2020
Am Hangfuß mündet unser Weg in einen breiteren Weg 4. Hier stehen wegnah sehr große Exemplare der amerikanischen Rot-Eiche (Quercus rubra) . Ihre spitz ausgezogenen Blattlappen kennzeichnen sie als „Spitzeiche“. Diese artenreiche Gruppe der Eichen (Sektion Lobatae) ist ursprünglich auf Amerika beschränkt, einige Arten werden aber heute an vielen anderen Orten angebaut. Die Rot-Eichen hier zeigen eine sehr üppige Naturverjüngung. Im Gegensatz zu unseren einheimischen Eichenarten benötigen die Eicheln der Rot-Eiche zum Reifen zwei Jahre.
Ein zweiter nordamerikanischer Baum, der seit dem 19. Jahrhundert auch in Mitteleuropa forstlich genutzt wird, ist die Douglasie oder Douglastanne (Pseudotsuga menziesii). Ob er wirklich die durch Klimawandel bedrohten Fichten als Forstbaum ersetzen kann, ist umstritten. Unbestritten ist seine große Wuchskraft. Der höchste Baum Deutschlands ist eine über 100 Jahre alte Douglasie im Arboretum Freiburg-Günterstal: „Waltraut vom Mühlwald“ ist über 100 Jahre alt und derzeit etwa 65 m hoch (https://www.waldhilfe.de/baumrekorde/?gclid=EAIaIQobChMIyaG0ztKY6wIVArp3Ch3YPwhKEAAYASAAEgJMZPD_BwE).
Douglasienzweig (Pseudotsuga mentziesii) mit typischen Zapfen. Im Unterschied zu Fichtenzapfen haben die Zapfen von Pseudotsuga sehr lange Deckschuppen, die aus den Zapfen heraushängen (Foto: W. Probst 1988)
Wir biegen dann wieder in eine halb links aufwärts führenden Waldweg ab. Von dort (5) folgen einige Teilnehmer*innen einem Trampelpfad bergauf, in der Hoffnung, auf den Ringwall zu stoßen. Wie die Rückkehrer berichten, konnten die Schanzenreste aber nicht eindeutig identifiziert werden. Am Wegrand wachsen zahlreiche Wald-Frauenfarne (Athyrium filix-femina). An einer Stelle kann man den zarteren Fiederschnitt ihrer Wedel mit einem Echten Wurmfarn (Dryopteris filix-mas) vergleichen. Die wissenschaftlichen Namen und die deutsche Bezeichnung „Frauenfarn“ weisen darauf hin, dass die beiden Arten früher für Männchen und Weibchen einer Art gehalten wurden.
Bei dem Silphienfeld stoßen wir wieder auf unseren alten Weg, der uns in wenigen Schritten zum Parkplatz zurückführt.
Während unseres ganzen Spazierganges durch den Wald hören wir die Rufe des Kolkrabe. Zum Schluss sehen wir ihn auf einer abgestorbenen Lärche sitzend (Foto: J. Probst, 8.8.2020)
Liste der demonstrierten Pflanzenarten in chronologischer Reiherfolge
zusammengestellt von der Exkursionsteilnehmerin Kim Hackenberg
„Mir ist die Anwesenheit von Anhängern der sozialistischen Sekte in unserer Provinz gemeldet worden, welche, mit geheimnisvollen Mixturen und übelriechenden Salben versehen, unsere arbeitsame Bevölkerung infizieren. Mit winzig kleinen und äußerst zerbrechlichen Ampullen ausgestattet, haben dieselben in Favara bereits eine starke und weit verbreitete Grippe ausgelöst (… …), und zwar mittels der Verbreitung von Keimen, die eine Mundschwamm-Epidemie auslösen können. Ich mache Sie darauf aufmerksam, daß diese Keime überaus leicht erkennbar sind: sie sind von leuchtend roter Farbe, jeder von ihnen besitzt 2.402 Füßchen. Für ihre Vernichtung muß Sorge getragen werden, weil sie sich außerordentlich schnell vermehren. In der Gewißheit, daß Sie sich der Gefahr bewußt sind und alle Maßnahmen für ein Einschreiten treffen, fordere ich Sie auf: gehen Sie ans Werk! S. E. der Präfekt (Vittorio Marascianno)“
Das schrieb der sizilianische Schriftsteller und Regisseur Andrea Camilleri in seinem 1998 erschienenen Roman „La concessione del telefono“; in deutscher Übersetzung 1999 beim Klaus Wagenbach Verlag mit dem Titel „Der unschickliche Antrag“ herausgegeben. Man könnte fast meinen, Camilleri hätte dabei an die Coronaviren gedacht. Das ist nicht ganz unmöglich, denn man kennt sie seit 1960. Der Roman allerdings spielt im Sizilien des ausgehenden 19. Jahrhunderts.
Allerdings konnte Camilleri nicht ahnen, dass die Corona Pandemie gute 20 Jahre später die Weltöffentlichkeit beherrschen würde, wie keine Epidemie, ja wie kein katastrophales Ereignis zuvor. Selbst die Klimakrise tritt dagegen zurück. Sie hat bisher bei weitem nicht so viele gleichlaufende Reaktionen in fast allen Ländern der Erde bewirkt, obwohl die Folgen vermutlich deutlich katastrophaler sein werden.
Coronaviren
Coronaviren haben einen Durchmesser von ca. 125 nm. Sie sind von einer mit zahlreichen Fortsätzen (Spikes) besetzten Proteinlipidmembran umgeben, an denen die Andockstellen für die entsprechenden Proteine der Wirtszellen liegen. Das genetische Material ist eine einsträngige RNA.
Coronaviren, kugeligen Gebilde mit den vielen Fortsätzen (Spikes), die sie im elektronenmikroskopischen Bild wie eine Sonne mit Corona erscheinen lassen, sind seit den 1960er Jahren als Erreger von Tierkrankheiten aus Großbritannien und den USA bekannt. 1968 erhielten sie ihren Namen. Aber erst 2003 wurde mit der SARS-Epidemie (severe acute respiratory syndrom) deutlich, dass sie auch für Menschen wirklich gefährlich werden können.
Unter den für menschliche Krankheiten verantwortlichen Viren gehören Coronaviren mit ihren 125 nm Durchmesser zu den größten. Mit 30.000 Nukleotiden ist ihre einsträngige RNA auch länger als die anderer RNA-Viren. Ein besonderer Reparaturmechanismus schützt diese lange RNA-Kette vor zu vielen falschen Replikationen.
Von Fledermäusen oder von Schuppentieren?
Wie andere Viren können Corona-Viren rekombinieren, wenn unterschiedliche Viren in einer Wirtszelle zusammentreffen. Aber eine Besonderheit der Corona-Viren, die sie besonders gefährlich macht ist, dass es nicht selten vorkommt, dass auch sehr unterschiedliche, genetisch weit voneinander entfernte Corona-Viren in einer Zelle zusammenkommen können und dass dann sehr gefährliche Rekombinanten entstehen. Besonders berüchtigt als Quelle solcher neuen Mischungen sind Fledermäuse, bei denen in einer Art bis zu zwölf verschiedene Coronaviren nachgewiesen werden konnten (Luis et.al 2013 nach Cyranoski 2020). Dabei haben die Fledermäuse offensichtlich einen Mechanismus entwickelt, der sie vor dem Ausbruch durch diese Viren hervorgerufener gefährlicher Krankheiten schützt.
Die zwei bisher aufgetretenen für den Menschen wirklich gefährlichen Coronaviren – SARS-CoV und MERS-CoV (middle east respiratory syndrom) – kommen beide von Fledermäusen. Deshalb liegt die Vermutung nahe, dass auch SARS-CoV2, der Erreger der derzeitigen Pandemie, von Fledermäusen abstammt, zumal es zwischen einem Fledermausvirus und SARS-CoV2 eine 96-%ige genetische Übereinstimmung gibt. Diese naheliegende Vermutung hat jedoch einen Haken: den Coronaviren in Fledermäusen fehlt allen an ihren Spikes eine besondere Proteineinheit (s.u.!), welche den Eintritt in menschliche Schleimhautzellen extrem erleichtert. Diese Proteinstruktur kommt aber bei Coronaviren von Schuppentiere (Fam. Manidae) vor. Die genetische Übereinstimmung des Schuppentier-Virus mit SARS-CoV2 beträgt jedoch nur 90 %. Neuere Arbeiten legen die Vermutung nahe, dass die virulenten Erreger schon vor 140 Jahren von Fledermäusen auf Schuppentiere wechselten. Während sie bei den Fledermäusen die gefährliche Proteinstruktur verloren haben, blieb diese beim Schuppentier erhalten (Cyranoski 2020).
Die unterschiedlichen Krankheitsbilder von Covid 19
SARS-CoV Viren infizieren die Schleimhäute von Nasenraum und Rachen als auch die des tieferen Lungengewebes. Im ersten Fall kommt es zu leichten bis mittelschweren Erkältungserscheinungen, im zweiten Fall kann die Erkrankung tödlich verlaufen. Auch Patienten, die zunächst an der leichten Form erkranken, können später noch eine gefährliche Lungeninfektion bekommen, und zwar dann, wenn ihr Immunsystem nicht bei der ersten Infektion Abwehrmechanismen entwickelt, die eine weitere Infektion verhindern. SARS-CoV und MERS-CoV können Rachen- und Nasenschleimhäute nicht infizieren. Mit der Infektion von Nasen- und Rachenschleimhäuten hängt auch zusammen, dass SARS-CoV2 Viren so leicht durch Tröpfcheninfektion übertragen werden können.
Die Speicheltröpfchen sind in Wirklichkeit – zum größten Teil – viel kleiner.
In den Speicheltröpfchen Infizierter befinden sich SARS CoV2-Viren
Ein besonderer Türöffner
Neben der leichten Übertragung durch Tröpfcheninfektion aus den oberen Atemwegen einer infizierten Person macht die Viren besonders gefährlich, dass sie die Membran der Schleimhautzellen sehr leicht überwinden und ihren Inhalt in diese Zellen befördern können. Dafür verantwortlich ist einmal die 8-10 mal höhere Affinität der Spikes zu der Andockstelle (Angiotensin converting enzym 2 ACE2, Wrapp et al. 2020), zum anderen eine besondere Proteinstruktur auf den Spikes, die nicht nur – wie bei den anderen Coronaviren – ein Andocken an bestimmte Proteine der Zellmembranen ermöglicht, sondern auch mit einem weiteren weitverbreiteten Enzym der Zelloberflächen von Epithelzellen, dem Furin, reagiert. Diese Protease spaltet den Kopf der Spikes von der restlichen Struktur ab und ermöglicht dadurch die Freisetzung von Fusionspeptiden, mit deren Hilfe die Proteinlipidmembranen von Virus und Epithelzelle verbunden werden. Ist das Virus erst einmal an eine Zelle angedockt, so ist es sehr wahrscheinlich, dass es auch seinem Inhalt in die Zelle transportieren kann. Wie diese Fusion auf molekularer Ebene genau funktioniert, ist allerdings noch nicht bekannt.
Nach Andocken des Virus an die Wirtszelle spalten Furine den Kopf der Spikes ab und setzen dadurch Fusionspolypeptide frei. Die Fusionspeptide bewirken eine Verbindung von Virushülle und Wirtszellmembran. Durch die Verbindung von Virus und Wirtszelle wird die Virus-RNA übertragen.
Mithilfe der Syntheseapparate der Wirtzelle werden die Bestandteile des Virus gebildet und zu neuen Viruspartikeln zusammengesetzt, welche die Wirtszelle verlassen und neue Zellen inizieren können.
Thrombosen und Organschädigungen
Studien aus den Niederlanden und Frankreich weisen darauf hin, dass 20-30 % der anCovid19 erkrankten Patienten Thrombosen entwickeln. Dabei handelt es sich zum Teil um sehr kleindimensionierte Thrombosen in Kapillaren. Eine mögliche Erklärung wäre, dass die Viren Zellen des Gefäßepithels befallen, die auf ihrer Zellmembran ebenfalls ACE2 und Furin tragen. Dadurch könnten die Gefäßinnenwände rauer werden und dies könnte die Bildung von Blutklümpchen begünstigen. Aber auch Effekte der Viren auf das Immunsystem könnten für die Thrombosen verantwortlich sein. Nachgewiesen ist, dass die Corvid19 Viren das Komplementsystem, also die unspezifische Immunantwort, in Gang setzen. Schädigungen von Organen wie Nieren, Leber, Herz und Nervensystem, die ebenfalls mit Corvid19 Infektionen in Verbindung gebracht werden, könnten auch eine Folge solcher Gefäßschädigungen sein.
Impfungen
Als der Erreger der Covid 19 Erkrankung zu Beginn des Jahres 2020 als SARS-CoV 2 identifiziert wurde, begann sofort an vielen verschiedenen Stellen die Entwicklung von und die Suche nach geeigneten Impfstoffen. Nach der Weltgesundheitsorganisation gibt es derzeit (2.Juni 2020) 148 solcher Impfstoffprojekte.
Aktive Schutzimpfung
Impfung mit inaktivierten (attenuierten) SARS-CoV-2 Viren
Impfung mit gentechnisch modifizierten Viren, die bereits für andere bewährte Impfstoffe verwendet wurden und bei denen man deswegen unerwünschte Nebenwirkungen ausschließen kann, zum Beispiel Vaccinia-Viren (bisher in Impfstoffen gegen Pocken), humane Adeno-Viren oder Masernviren.
Impfung mit aus ausgewählten Genen des Virus in Form von mRNA oder DNA. Nach Injektion in Körperzellen sollen sie diese zur Bildung von ungefährlichen Virusproteinen anregen, die dann den Aufbau des körpereigenen Immunschutzes bewirken. So enthält mRNA-1273 die Erbinformation für einen Bestandteil des Spike-Proteins von SARS-CoV-2. Der Impfstoff bewirkt in Körperzellen die Biosynthese von diesem Protein, das als Antigen wirkt und das körpereigene Immunsystem zur Bildung von Antikörpern gegen das Virus anregt .
Passive Schutzimpfung
Impfung mit Immunglobulinkonzentraten von Personen, die eine Covid 19 Erkrankung überstanden haben. Werden die aus dem Serum isolierten und gereinigten Antikörper einem anderen Coronapatienten injiziert, so erhält er eine „passive Immunisierung“. Diese Impfung wird deshalb teilweise auch als Serum-Therapie bezeichnet. Die übertragenen Antikörper können dabei sofort gegen die Krankheitserreger wirksam werden.
Die Serum-Therapie hat bei der Bekämpfung der Ebola-Epidemie bereits gute Dienste geleistet. Durch ihren Einsatz konnte die Sterblichkeitsrate bei dem Ebolaausbruch im August 2018 in der Demokratischen Republik Kongo um 30 % reduziert werden.
Die passive Schutzimpfung mit Antikörpern aus genesenen Corona-Patienten kommt vor allem für Risikogruppen und für schwer erkrankte Patienten infrage. Sie ist kein Ersatz für eine langfristig wirkende aktive Schutzimpfung, denn sie wirkt in der Regel nur wenige Wochen bis Monate. Es entsteht keine bleibende Immunität gegen den Erreger, da die künstlich zugeführten Antikörper innerhalb von etwa 30 Tagen wieder abgebaut werden.
Spekulationen
Der besonderer Türöffner an den Spikes von SARS-CoV2 ist ein Grund dafür, dass immer wieder spekuliert wird, der besonders gefährliche Keim wäre in einem Labor gezielt hergestellt und absichtlich oder unabsichtlich freigesetzt worden. Dafür gibt es allerdings bisher keinerlei Anhaltspunkte. Aber alleine die Tatsache, dass in Wuhan, der chinesischen Millionenstadt, von der die Epidemie ihren Ausgang nahm, am Wuhan Institute of Virologie seit der SARS-Epedemie von 2003 intesiv an Coronaviren georscht wird, genügt Verschwörungstheoretikern, um hier einen Zusammenhang herzustellen.
Dabei wird auch dem Microsoft-Milliardär Bill Gates, der schon lange vor dem möglichen Ausbruch einer weltumspannenden Pandemie gewarnt hat, eine besonders finstere Rolle zugedacht. Schon vor zehn Jahre kommentierte Gates den überstandenen H1N1-Ausbruch von 2009 mit den Worten: „Wir hatten Glück, dass es nicht schlimmer kam. Denn wir waren fast komplett unvorbereitet.“ Im folgenden wies er in Reden und Artikeln immer wieder auf die Pandemie-Gefahr hin: „Wenn irgendetwas in den nächsten Jahrzehnten mehr als zehn Millionen Menschen tötet, ist das wahrscheinlich ein Virus und nicht ein Krieg“.(2015; nach NZZ 11.04.2020). Diese weise Voraussicht, die sich im Nachhinein als richtig erwies, dient nun Verschwörungstheoretiker dazu, Gates finstere Komplotte zur Errichtung einer Weltdiktatur zu unterstellen. Hat er gar mit chinesischen Virologen zusammengearbeitet mit dem Ziel, als Folge der Maßnahmen gegen die Pandemie eine totale Kontrolle über die Menschheit zu gewinnen? Auch seine Unterstützung der World Health Organisation – nach Ausscheiden der USA mittlerweile der größte Einzelunterstützer – wird in diese Richtung interpretiert.
Im Prinzip ist es tatsächlich problematisch, wenn internationale und nationale staatliche Aufgaben zunehmend von der Unterstützung einzelner Milliardäre abhängig werden. Deshalb ist es sehr schade, dass US-Präsident Trump die Vereinigten Staaten aus der WHO herausnehmen und diese Organisation nicht mehr unterstützen will. Bedenklich ist auch, dass er dies mit ähnlichen Argumenten begründet, wie die Verschwörungstheoretiker: Nachdem er die Maßnahmen Chinas gegen die Coronaepidemie zunächst gelobt hatte, bezichtigt er China nun einer bewussten Täuschung der Weltöffentlichkeit. Der WHO wirft er eine chinafreundliche Politik vor, wohl vor allem, um damit Fehler der eigenen Politik in Sachen Corona zu vertuschen.
Andere Spekulationen, die von allen möglichen „Weltverbesserern“ und Ideologen aufgestellt werden, sind noch wesentlich abstruser. In der TAZ schreibt Arno Frank dazu: „Gefährlich und grotesk wird es, wenn das blinde Huhn beim Stochern drei Körner findet und daraus ein Vollkornbrot backen will:“ (TAZ 23./24.5.2020, S.3).
Weltweite Kooperation
Auch wenn die unter den Begriff „Lock Down“ in vielen Ländern der Erde getroffenen Maßnahmen zur Minderung der Infektionsgefahr nicht immer sinnvoll und manchmal auch überzogen erscheinen mögen, so zeigen sie doch, dass die Menschheit angesichts drohender Gefahren zu gleichsinnigem Handeln in der Lage ist. Es besteht deshalb eine gewisse Hoffnung, dass diese Erfahrungen sich auch auf die Handlungsfähigkeit Hinblick auf den Klimawandel positiv auswirken könnten. Eine erste Chance ergibt sich hier schon bei den gezielten Maßnahmen zur Wirtschaftsförderung. Im Hinblick auf Klimaziele geforderten Veränderungen der Wirtschaftssysteme könnten in der gegenwärtigen Situation durch gerichtete Förderung leichter in Angriff genommen werden.
Quellen
Cyranoski, D. (2020): Profile of a killer virus. Nature 581, pp. 22-26
Probst, W., Schuchardt, P. (Hrsg., 2020): Basiswissen Schule Biologie – Abitur. 5. Auflage. Berlin: Duden
Willyard, C. (2020): Coronavirus blood-clot mystery intensifies. Research begins to pick apart the mechanisms behind a deadly COVID-19 complication. Nature 581, p. 250
Wrapp, D. et al. (2020): Cryo-EM structure of the 2019-nCoV spike in the prefusion conformation.Science 367 (6483), pp.1260-1263
Die Erde bildete sich vor etwa 4,6 Mrd. Jahren. 0,5 bis 1 Mrd. Jahre später traten die ersten Lebewesen auf und schon vor ca.3,5 Mrd. Jahren entwickelten sich die ersten Cyanobakterien, die mithilfe von Sonnenlicht aus Wasser und Kohlenstoffdioxid Kohlenhydrate und Sauerstoff herstellen konnten. Der Sauerstoff oxidierte Mineralien und löste sich in den Ozeanen. Erst nach etwa 1 Mrd. Jahren waren diese Oxidationsprozesse abgeschlossen und der O2-Gehalt der Atmosphäre begann stark anzusteigen – mit tödlichen Folgen für obligate Anaerobier aber mit einem großen Vorteil für Lebewesen, die zur aeroben Atmung mit Sauerstoff in der Lage waren. Mit Photosynthese und Atmung war die Grundlage für effektive chemische Kreisläufe in der Biosphäre geschaffen.
Seither hat sich die Stoffproduktion durch Photosynthese stetig vermehrt, auch wenn es immer wieder kleinere oder größere Rückschritte gab. Vor etwa 400 Mio J. begann die Besiedelung des Festlandes durch grüne Pflanzen und dieser grüne Pelz überzog von Feuchtgebieten ausgehend immer größere Flächen der Kontinente. Der Pelz wurde auch immer dichter und höher. Die höchsten Bäume können über 100 m hoch werden und die Pflanzendecke ist vielfach geschichtet. Die Pflanzen wurden durch natürliche Selektion an immer extremere Lebensbedingungen angepasst, sodass immer trockenere und immer kältere Gebiete einen grünen Pelz bekamen.
Beschädigungen
Waren in der früheren Erdgeschichte vor allem plattentektonisch bedingte Veränderungen der Kontinente, Vulkanausbrüche und Asteroideneinschläge aber auch biogene Veränderungen des CO2-Gehalts der Atmosphäre für Rückschritte bei dieser Entwicklung verantwortlich, so ist es heute die menschliche Zivilisation, durch die der grüne Pelz des Bioplaneten Erde beschädigt wird. Diese Beschädigungen haben mittlerweile ein Stadium erreicht, das für die menschliche Zivilisation und für die derzeitigen Ökosysteme gefährlich wird. Denn angesichts der großen Populationsdichte der Menschen und des Zivilisationsgrads wird der grüne Pelz der Erde verringert und in seiner Wirksamkeit beeinträchtigt.77% der Landfläche (ohne Antarctica) und 87 % der Meere sind derzeit durch menschliche Aktivitäten verändert worden (Watson, Allen u.a.2018).
Städte werden immer größer, Verkehrsnetze immer
dichter, Agrarflächen, die mit ihren Monokulturen eine deutlich geringere
regulatorische Wirkung haben als natürliche Vegetation, dehnen sich immer
weiter aus und lassen das grüne Fell der Erde räudig werden.
Die Kapazität des grünen Pelzes wird im Hinblick
auf eine ausgeglichene Stoffbilanz des Bioplaneten Erde dadurch überschritten,
dass fossile Energieträger zur Energiebereitstellung verbrannt und zur
(Kunst-)Stoffproduktion genutzt werden. Besonders die starke Zunahme des
Treibhausgases CO2 führt zu einer deutlichen Klimaerwärmung.
Der Eingriff in den Stickstoffkreislauf durch
anthropogene Umwandlung des Luftstickstoffs (N2) in
reaktionsfreudige Stickstoffverbindungen kann sich über verminderte
Biodiversität und Veränderung der Atmosphäre (Verringerung der UV-Licht
filternden Ozonschicht) negativ auswirken.
Diese Veränderungen stellen für den Bioplaneten keine existentielle Gefahr dar, das Leben auf der Erde wird diese Veränderungen ebenso überstehen, wie es andere oft noch viel drastischere Ereignisse im Laufe der Erdgeschichte überstanden hat. Für die menschliche Zivilisation in ihrer heutigen Form stellen sie aber eine existentielle Bedrohung dar. Für eine nachhaltige Entwicklung des Bioplaneten als Lebensraum für die Menschen ist der Erhalt des grünen Pelzes deshalb von entscheidender Bedeutung.
Städte
Sao Paulo,12,3 Mio Einwohner (Quelle: pixibay, joelfotos)
Mit der zunehmenden Bevölkerung werden Städte immer größer und überdecken immer größere Flächen (Liu u.a.2020). Herkömmliche Städte sind nicht grün, sie haben Oberflächen, die vorwiegend aus Beton, Steinen, Glas und Asphalt bestehen. Die photosynthetische Stoffproduktion ist niedrig, die CO2-Produktion ist viel höher als der CO2-Verbrauch, C-Speicherug in Vegetation und Boden ist gering. Ebenso gering im Vergleich zu natürlichen Ökosystemen ist das Rückhaltevermögen für Regenwasser, sodass es bei den durch Klimawandel vermehrten Starkregen immer häufiger zu Überschwemmungen kommt. Pflanzliche Oberflächen verdunsten Wasser und produzieren Verdunstungskälte. Steine und Beton speichern Wärme. Beides führt dazu, dass das Stadtklima wärmer ist als das Klima in der Umgebung. Dabei spielt auch eine Rolle, dass der Luftaustausch mit der Umgebung durch die Gebäude behindert wird.
Eine Stadt mit großen Grünanlagen wie Parks und Gärten bietet zwar eine hohe Lebensqualität und eine bessere Ökobilanz. Dies geht aber insofern auf Kosten der Umgebung, als sie mehr Fläche für denselben umbauten Raum benötigt. Wenn die Umgebung aus intensiv bewirtschafteten Ackerflächen besteht, kann deren Umwandlung in gartenreiche Wohngebiete trotzdem Vorteile bieten (Reichholf 2018). Für die heutigen, von dicht stehenden Hochhäusern dominierten Großstädte ist das aber keine realistische Alternative, da die benötigten Flächen viel zu groß wären. Eine Erfolg versprechende Möglichkeit für dicht bebaute Großstädte ist die Integration von Bauwerken und Grünanlagen.
Schon lange zählt es zu Attributen ökologischer Bauweise, Dächer zu begrünen. Die Etablierung und Ausgestaltung solcher Dachgärten und Wiesen ist aber noch sehr stark ausbaufähig, wie man auf Luftbildern von Städten leicht erkennen kann. Neben der Flächenvergrößerung könnte auch die Ausgestaltung verbessert werden. Dickere Bodenschichten verbessern die Stoffbilanz, die Wasser- und Kohlenstoff-Speicherung. Zisternensysteme können für die Bewässerung während Trockenperioden genutzt werden und den Wasserabfluss bei Starkregen mindern.
Begrünte Dachflächen könntemn durch Brücken verbunden werden.
Vernetzte Dachgärten (Entwurf W. Probst, 2020)
Auch begrünte Fassaden gibt es schon lange, aber eher an alten Bauernhäuser auf dem Land als an mehrgeschossigen Stadthäusern, Bankhochhäusern und Industrieanlagen. Für diese traditionelle Fassadenbegrünung sind vor allem Lianen wie Efeu oder Wilder Wein (Parthenocissus) verantwortlich, die sich mit besonderen Haftorganen an den Fassaden festhalten – ein Grund dafür, dass sich viele Hausbesitzer wegen der dadurch erschwerten Fassadenrenovierung davon abhalten lassen, eine solche Wandbegrünung zu erlauben. Auch die Furcht vor Beschädigungen durch die wuchernden, oft auch in Risse und Öffnungen eindringenden Lianen spielt dabei eine Rolle. Diese Probleme können durch vorgebaute Rankgerüste teilweise vermindert werden. Eine staatlich finanzierte Förderung der Fassadenbegrünung, wie sie ähnlich bei Fassadendämmungen sehr erfolgreich angewendet wird, könnten ein wirkungsvoller Anschub sein. Besonders wirkungsvoll könnte eine solche Förderung werden, wenn flächenhafte Begrünungsmodule zur Verfügung stünden, die mit einfachen Mitteln an Fassaden angebracht werden könnten und die durch Anschluss an eine Bewässerungsanlage wartungsarm wären. Die Elemente könnten aus einem Gerüst bestehen, an dem mehrere auswechselbare Pflanzgefäße aufgehängt werden.
Eine weitere Möglichkeit der vertikalen Begrünung, die in wenigen Beispielen schon verwirklicht ist, wäre die Ausgestaltung von Pflanzbalkonen mit Sträuchern und Bäumen. Große Gebäudekomplexe könnten durch grüne Brücken vernetzt werden. Verkehrswege, insbesondere Straßen und Schienenverkehr, könnten wie U-Bahnen unter die Oberfläche verlegt werden, wodurch Platz für bodenständige Grünanlagen aber auch Rad- und Fußwege gewonnen würde, So könnten schließlich Städte entstehen, die ganz in einem grünen Pelz eingehüllt sind und die sich fast übergangslos in die umgebende Landschaft einfügen (vgl. Jean Nouvel 2014, Stefano Boeri 2015).
Verkehrswege
Verkehrswege, insbesondere Straßen für den KFZ-Verkehr, tragen einmal durch Versiegelung zur Reduktion des grünen Pelzes bei, zum anderen zerschneiden sie Ökosysteme, führen zur Verinselung und darüber insbesondere zur Schädigung von Tierpopulationen und damit zur Verringerung der Biodiversität. Schließlich belasten die Abgase der Kraftfahrzeuge die Luft.
Autobahn (Quelle: pixabay: Alexas_Fotos)
Mögliche Verbesserungen:
Stichworte
Zerschneidungseffekte minimieren (Brücken über wertvolle Landschaftsteile, grüne Brücken zur Minderung von Zerschneidungseffekten, Tunnel),
Verkehrsflächen unter die Erde verlegen; nicht nur Hindernisse (Berge, Gewässer) sondern auch besonders schützenswerte Landschaften untertunneln,
emissionsarme Verkehrsmittel nutzen.
Je dichter die Besiedelung, desto dichter sind nicht nur Städte,
Siedlungen und Industrieanlagen, desto
dichter ist auch das Netz von Verkehrswegen, insbesondere Straßen und
Autobahnen (in Deutschland derzeit nach
Erhebung des Umweltbundesamt knapp 20000 km², das entspricht rund 5,5% der Landesfläche). Das wirkt sich aber nicht nur
über den Flächenverbrauch sondern vor allem über den Zerschneidungseffekt
nachteilig auf die Funktion von Ökosystemen aus. Mehr noch als Pflanzenarten
sind Tierpopulationen durch die dadurch bedingte Verinselung betroffen. Auch
die direkte Tötung von Tieren durch den Verkehr spielt eine Rolle. Indirekt
wirkt sich dies über die Bestäuber und die Verbreitung von Früchten und Samen
auf die Vegetation aus.
Eine Verbesserung kann einmal durch geeignetes Straßenbegleitgrün erreicht werden (Kühne/Freier 2012). Vor allem aber kann die trennende Wirkung von Verkehrsflächen durch Brücken, sowohl Brücken über schützenswerte Landschaftsteile als auch verbindende Grünbrücken, und Tunnel erreicht werden. Schutzgräben oder Zäune können in Kombination mit kleinen Tunneln insbesondere Amphibien bei ihren Laichwanderungen schützen (Krötenzaun, Krötentunnel).
Natürlich ist das Hauptproblem die hohe Verkehrsdichte und die Emissionen der Verkehrsmittel. Sie wird einmal durch den Individualverkehr, zum anderen durch den Güterverkehr verursacht. Beide haben in den letzten Jahrzehnten ständig zugenommen. Eine größere Verlagerung dieses Verkehrs auf die Bahn wird schon lange als Ziel formuliert, ließ sich aber bisher politisch nicht durchsetzen. Auch eine Förderung dezentraler Produktion könnte der ständigen Zunahme des Güterverkehrs entgegenwirken.
Landwirtschaft/Nahrungsmittelerzeugung
Moderne Landbewirtschaftung hat zwar zu immer höheren Erträgen pro
genutzter Fläche geführt, die Gesamtstoffbilanz, in die man den Verbrauch von
fossilen Energieträgern einrechnet, ist aber immer schlechter geworden. Nach
Smil (2019) wird heute pro Ackerfläche 10x soviel produziert wie vor 100 Jahren
aber dafür wird 90x soviel Energiezufuhr benötigt.
Riesige Monokulturen, Pestizid- und Düngemitteleinsatz erhöhen zwar die landwirtschaftliche Produktion, vermindern aber insgesamt die Leistungsfähigkeit des grünen Pelzes und schädigen Böden und ihre Kohlenstoff-Speicherfähigkeit. Artenarme, mit Pestiziden behandelte Agrarflächen sind die Hauptursache für den starken Rückgang der biologischen Vielfalt. Die Massentierhaltung ist nicht nur ein ökologisches sondern auch ein ethisches Problem.
Weizenfeld nach der Ernte (Quelle: pixabay: ulleo)
Feldgehölze und andere artenreiche Biotope als
ökologische Inseln
Agroforestry
Vertical Farming
Landwirtschaft 4.0 (KI)
Das gewichtigste Argument für eine immer stärkere Rationalisierung und Industrialisierung der Landwirtschaft ist, dass nur dieser Weg für die ständig steigenden Bedürfnisse der wachsenden Erdbevölkerung die notwendigen Nahrungsmittel und weiteren Rohstoffe liefern kann. Dieses Argument greift aber insofern nicht, als die derzeitige Landbewirtschaftung auf irreversiblem Verbrauch basiert, Verbrauch von fossilen Energieträgern, Verbrauch von Wasser, Verbrauch von nicht regenerierbaren Düngemitteln (insbesondere Phosphat, Greuling 2011), Verbrauch von Böden, Verbrauch von selbstregulierenden Ökosystemen wie z.B. Regenwäldern.
Systeme, die auf Verbrauch basieren, sind aber nur nachhaltig, das
heißt, für längere Zeit funktionsfähig, wenn die verbrauchten Ressourcen
ständig regeneriert werden können, Dies ist gegenwärtig eindeutig nicht der
Fall. Deshalb ist eine Veränderung
vorhersehbar. Sie kann nur ohne Katastrophen stattfinden, wenn sie basierend auf wissenschaftlichen
Erkenntnissen der Ökologie vorgenommen wird.
Das kann natürlich nicht bedeuten, dass man zu Methoden des
Neolithikums zurückkehrt. Eine den Produktionserfordernissen der Gegenwart
genügende Landbewirtschaftung, die gleichzeitig nachhaltig ist, bedeutet nicht
weniger Technik sonder mehr Technik, genauer gesagt mehr intelligente Technik.
Sehr große, von Monokulturen bestandene Flächen erlauben den Einsatz von riesigen Maschinen und haben dazu geführt, dass mit wenigen menschlichen Arbeitskräften große Stoffmengen produziert werden können. Gleichzeitig werden dadurch aber lebenswichtige Ressourcen, Artenvielfalt, Böden, Dünger und Energie liefernde Stoffe „verbraucht“ und andere Ökosysteme durch Eintrag von Düngemitteln und Schadstoffen geschädigt.
Das Grüne Band Deutschland bezeichnet einen Geländestreifen entlang der ehemaligen innerdeutschen Grenze, der als arten- und biotopreicher Grüngürtel erhalten bleiben soll und der zudem wertvolle Biotope miteinander verbindet. Wenn von diesem grünen Band weitere Grüngürtel ausgehen würden, könnte es Ausgangspunkt für eine landesweite oder sogar europaweite Netzstruktur werden.
Würden die Monokulturen durch ein Netz naturnaher linearer Elemente wie Feldhecken und Wildpflanzenstreifen unterbrochen, könnte dieser Verbrauch zwar gemindert werden, gleichzeitig wäre aber eine Bewirtschaftung mit den derzeit üblichen Methoden nicht möglich oder viel aufwändiger. Mit kleineren, intelligenten Maschinen, wie sie in einfacher Form heute schon allgemein zum Staubsaugen oder Rasenmähen eingesetzt werden, wäre das aber durchaus denkbar. Solche intelligenten, lernfähigen Roboter könnten – mit Luftbildern von Drohnen oder auch Satelliten versorgt – sehr gezielt arbeiten. Zusammen mit der Roboter eigenen Sensorik würde eine gezielte und damit sparsamere Unkrautvernichtung, Schädlingsbekämpfung, Düngung und Bewässerung möglich. Statt flächendeckender Düngung könnten gezielt nur solche Teilbereiche gedüngt werden, die tatsächlich unterversorgt sind. Pestizide könnten nur auf tatsächlich befallene Pflanzen gesprüht werden, dasselbe gilt für die Bekämpfung von Unkräutern. Statt Riesentraktoren und Megamaschinen würden dann viele kleine Roboter die Ackerflächen bearbeiten. Eine solche von künstlicher Intelligenz bestimmte Agrarwirtschaft wird auch als Landwirtschaft 4.0 bezeichnet.
Alternative, Ressourcen schonendere Formen der Landbewirtschaftung wie
Mischkulturen und Agroforestry, spielen heute nur in Nischen und
Subsistenzwirtschaften eine Rolle, da sie sehr arbeitsintensiv sind. Durch
Einsatz intelligenter Technik könnten manuelle Tätigkeiten durch Roboter und
Regelsysteme ersetzt und damit solche nachhaltigen Wirtschaftsformen rentabler
werden.
Eine weitere zukunftsweisende Form zur Produktion von Nahrungsmitteln und anderen nachwachsenden Rohstoffen wird mit dem Begriff „Vertical Farming“ bezeichnet. Dadurch könnte der Flächenverbrauch der Produktion stark verringert werden. Schon auf der Internationalen Gartenschau in Wien 1964 wurde ein von dem Maschinenbauingenieur Othmar Ruthner konstruiertes Turmgewächshaus gezeigt. Weitere Verbreitung dieser Idee sorgte der New Yorker Professor für Umweltgesundheit und Mikrobiologie Dickson Despommier, der mit seinen Studenten ab 1999 entsprechende Ideen zunächst für die Nahrungsmittelversorgung der 50000 Einwohner Manhattans entwickelte. Ausgangspunkt waren Überlegungen zum möglichen Gemüseanbau auf Dachflächen. In der Weiterentwicklung wurden Hochhäuser geplant, die insgesamt der Pflanzenkultur dienen sollen. In jedem Stockwerk eines solchen Hochhauses sollen Pflanzen auf optimale Weise automatisch gesteuert und reguliert kultiviert werden. Gleichzeitig sind diese Kulturen in Kreislaufsysteme, insbesondere der Wasserwiederverwendung und Abwasseraufbereitung, eingebunden (Despommier 2011).
Das Prinzip „Wachsen lassen“
Wenn die möglichst optimale
Förderung der Vegetation als wichtigstes Naturschutzziel im Sinne einer für die
menschliche Zivilisation nachhaltigen Entwicklung des Bioplaneten anerkannt
wird, müssen Pflanzenwachstum und Vegetationsentwicklung so gut wie möglich
gefördert werden. Das bedeutet, dass man Pflanzen überall dort wachsen lässt,
wo sie nicht wichtige Funktionsabläufe stören.
Die Bearbeitung von Kulturflächen ist in vielen Fällen notwendig. Wenn
man eine Wiese in Mitteleuropa nie mehr mäht, wird daraus in ein, zwei
Jahrzehnten ein Gebüsch und in einem Jahrhundert ein Hochwald. Einen Acker muss
man regelmäßig bestellen, abernten, düngen und auch spritzen, um ernten zu
können. Aber wie sieht es mit den Rändern und den Grenzen zwischen den
verschiedenen Nutzungsflächen aus? Hier besteht für den Naturschutz ein
riesiges Potenzial, das für den Naturhaushalt vermutlich ergiebiger ist, als
die in ihrem Flächenanteil sehr beschränkten Naturschutzgebiete. Außerdem hilft
der Randschutz, verinselte naturnahe Flächen zu vernetzen. Eine
vielversprechende Initiative, welche diese Idee verfolgt, ist das „Konzept der Ehda-Flächen“. Initiator und Träger dieses Projektes ist das
Institut für Agrarökologie des Landes Rheinland-Platz (IfA). In den Stadtkernen betrifft dies Parkanlagen,
aufgegebene Verkehrsflächen, Brachflächen, die vorübergehend nicht bebaut sind,
Randstreifen und Verkehrsinseln, die man
zeitweilig der Spontanvegetation überlassen kann. Auch die Grünflächen um
öffentliche Gebäude wie Krankenhäuser, Verwaltungs- und Regierungsgebäude
liefern große, bisher nicht sinnvoll genutzte Flächen.
Ein besonders großes Potenzial stellen Privatgärten dar, die meist in
den Randbereichen der Städte in Vierteln
mit Einfamilien- und Reihenhäusern konzentriert sind. Hier gilt meist das
Prinzip, dass nur wachsen darf , was gepflanzt wurde. „Un“kraut jäten ist
deshalb neben Rasen mähen und Hecken
schneiden die häufigste Beschäftigung des Hobbygärtners. Um das Unkraut ohne zu
viel manuelle Tätigkeit fern zu halten, hat sich schon vor einigen Jahrzehnten
verbreitet, die Beete mit einer Schicht aus keimungs- und wachstumshemmendem
Rindenmulch zu bedecken.Seit einigen Jahren wird eine noch pflanzenfeindlichere
Methode, das Auskiesen von Gartenflächen, immer beliebter.
Durch solche Maßnahmen gehen sehr viele potenzielle Flächen für einen
ökologisch wirkungsvollen „grünen Pelz“ verloren.
Einige Regeln, die helfen können, aus einem Garten eine ökologisch
wertvolle Grünfläche zu machen:
Zierpflanzen, die gut gedeihen, fördern, auf solche, die schlecht wachsen oder sehr viel Pflege benötigen, verzichten,
auf Pestizide verzichten oder sie nur sehr gezielt bei einzelnen befallenen Pflanzen einsetzen,
Wildpflanzen nur entfernen, wenn sie gewünschte Zier- oder Nutzpflanzen schädigen oder verdrängen,
Wildpfanzen unter Hecken oder Sträuchern wachsen lassen,
Rasenflächen, die rein ornamentale Funktion haben, zu mageren (nicht gedüngten), höchstens zweimal im Jahr gemähten Wiesen umwandeln,
Abstellflächen (z.B. Autostellplätze) nicht pflastern oder asphaltieren, sondern als Schotterrasen gestalten,
Einfahrten mit unterbrochenen Pflastersteinen befestigen, die Bewuchs und Wasserversickerung ermöglichen,
abgeblühte Blütenstände und abgestorbene Fruchtstände wenigstens teilweise stehen lassen, auch über Herbst und Winter (Überwinterungsplätze für Insekten)
Gartenabfälle vor Ort kompostieren,
aus Strauch- und Baumschnitt Reisighaufen anlegen,
Gartenmauern als Trockenmauern anlegen, Mauerritzen können zur schnelleren Begrünung mit passenden Pflanzen geimpft werden (Zimbelkraut, Mauerraute, Schöllkraut, Polster von Mauermoosen wie Drehzahnmoos, Kissenmoos)
Abwechslungsreiche Besiedelungsflächen schaffen (Sandflächen, Lehmböden, humusreiche Böden, Stein- bzw. Bauschutthaufen),
Regenwasser vom Dach (und versiegelten Flächen) in Zisterne sammeln und als Gießwasser (ggf. auch für Teich /Bachlauf) nutzen.
Wildwuchs an der Gartengrenze (Großblutige Königskerze – Verbascun densiflorum)
Quellen
Blanc.P. (2009): Vertikale Gärten, Die Natur in der Stadt. Stuttgart:
Ulmer
Boeri, S. (2015): A vertical Forest. Milano: Editione Mantova
Delwiche, C., F., Cooper, E., D. (2015): The evolutionary origin of terrestrial flora. Current Biology25, S. R899 – R919
Haft, J. (2.A. 2019): Die Wiese – Lockruf in eine
geheimnisvolle Welt. München: Pengiun
Hendershot, J., N. u.a. (2020): Intensive farming
drives long-term shifts in avian comunity composition. Nature 579, p.393-396
Kühne, S./Freier, B. (2012): Saumbiotope und ihre Bedeutung für Artenvielfalt und biologischen Pflanzenschutz. Workshop „Biological Diversity in Agricultural Landscapes“ – February 09-10, 2012, Berlin-Dahlem
Liu, Xiaoping et al. (2020): High spatiotemporal resolution mapping of global urban change from 1985 to 2015: Nature Sustainability. DOI: 10.1038/s41893-020-0521-x
Probst, W. ,Hrsg. (2017): Saumbiotope – Grenzen
und Übergänge. Unterricht Biologie 425. Seelze: Friedrich
Reichholf, J. H. (2018): Schmetterlinge: Warum sie verschwinden und was das für uns bedeutet. München: Hanser
Schilk, D. (2019): Die Wiederbegrünung der Welt. Klein Jasedow: Drachen-Verlag
Smil, V. (2019): Growth – From microorganismes to megacities. Cambridge MA.: MIT-Press
Watson, J. E. M., Allen, J. A. U:A: (2018): Protect the last of the wild. Nature 563, pp. 27-30
Mehl aus reifen Samen, unreife Bohnen geröstet oder
gekocht
Afrika
Trockengebiete Westafrikas
Bockshornklee (Trigonella foenum-graecum)
Samen als Nahrungsmittel und Tierfutter, Gewürz (z. B. in
türkischer Gewürzpaste Çemen), Sprossen (verursachten wg. bakterieller Verunreinigung EHEC-Epidemie),
medizinisch möglicherweise gegen Parkinsonsymptome (Verzögerung
der Schädigung dopinerger Nerven)
Süd- u. Mitteleuropa, Afrika, Naher Osten, Indien, China,
Australien
das harzige Fruchtfleisch der röhrenförmigen Hülsen wird ähnlich
wie Lakritz genutzt und ist Bestandteil von Abführmitteln; dient auch zum
automatisieren von Tabak
alle Pflanzenteile, auch die eiweißreichen Knollen sind
essbar, Ölgewinnung
Südostasien, Afrika (vor allem im Gartenbau)
Afrika, Madagaskar
Gartenbohne (Phaseolus vulgaris)
sehr viele Sorten
mit unterschiedlich gefärbten und geformten Samen
unreife Hülsen („grüne Bohnen“) und reife Samen als Gemüse
weltweit mit Schwerpunkt in Asien
Mittelamerika und andines Südamerika
Guarbohne (Cyamopsis tetragonobolus)
Hülsen als Gemüse und Grünfutter; Guarkernmehl mit
Hauptbestandteil des Mehrfachzuckers Guaran als Verdickungsmittel (E 412);
technisch für Fracking verwendet
Indien und Pakistan,
Südwest-USA, Australien, Israel
vermutlich Indien
Hamburger Bohne,
Samtbohne (Mucuna sloanei)
gekochte unreife Hülsen und reife Bohnen als Gemüse;abführend
und harntreibend;aus allen Pflanzenteilen kann ein schwarzer Farbstoff gewonnen
werden
grüne Hülsen und unreife Bohnen als Gemüse, reife Bohnen
als Gemüse oder Mehl, in Ägypten Gebäck „Tauniah“ (auch aus Vicia faba); Bodenverbesserung und
Gründüngung; in Ostasien medizinisch genutzt
junge Hülsen und unreife Bohnen als Gemüse, reife Samen
nach entwässern und langem Erhitzen als Gemüse
subtropische und tropische Gebiete weltweit
Mittel und Südamerika
Johannisbrot (Ceratonia siliqua)
Frucht wird frisch oder getrocknet verzehrt, zu Saft
gepresst, zu Sirup verarbeitet oder zu alkoholischem Getränk vergoren, das
getrocknete Fruchtfleisch wird auch zu Carobpulver vermahlen und wie
Kakaopulver genutzt; die Samen liefern ein technisch genutztes
Verdickungsmittel, Carubin oder Johannisbrotkernmehl (E 410)
Mittelmeergebiet, insbesondere Spanien einschließlich der
Balearen
Mittelmeergebiet und Vorderasien
Juckbohne (Mucunia pruriens)
in den Tropen weit verbreitete Futterpflanze (Heu,
Silage), Bohnen geröstet als Kaffeeersatz, als Gemüse nur nach langem
Einweichen und Kochen, da roh giftig (Alkaloide)
Heilpflanze, z. B. gegen Parkinson; Haare für Juckpulver
Mittelmeerländer, subtropische bis warm- gemäßigt Zonen
weltweit
Mittelmeergebiet
Mattenbohne, Mottenbohne
(Vigna aconitifolia)
ganze oder halbierte Bohnen als Gemüse gekocht oder
fritiert
Indien, Sri Lanka, asiatische Trockengebiete
Indien
Mungobohne ,
„Green Gram“ (Indien) (Vignaradiata)
ganze oder halbierte Bohnen als Gemüse gekocht oder
frittiert, Mehl für verschiedene Gerichte, z. B. Glasnudeln; grüne Bohnen als
Salat oder Gemüse, Sprossen
reife Bohnen vor allem als Viehfutter aber auch für die
menschliche Ernährung
Indien
Südindien
Platterbse, Saat-,
Kicherling, (Lathyrus sativus)
Mehl, Schrot, Gemüse
Europa (v.a. Mittelmeergebiet), W-Asien
Mittelasien (pontisch-kaspisch)
Reisbohne (Vigna
umbellata)
trockene Bohnen werden wir Reis zubereitet; grüne Hülsen
und junge Bohnen als Gemüse und Salat, Sprossen
Südostasien, Indien
Zentralchina bis Malaysia
Schabziger-Klee;
Brotklee (Trigonella caerulea)
Gewürz, v.a. zur Herstellung des Schabziger-Käses (Kräuterkäse,Schotterkäse,Grüner
Käse)
Alpenraum, v. a. Schweiz, Südtirol,
Ursprungsart T.
.procumbens im östlichen Mittelmeergebiet und Kaukasus
Schwertbohne (Canavalia
gladiata)
reife Körner müssen wie bei der Jackbohne entgiftet werden
subtropische und tropische Gebiete weltweit
Südasien und Afrika
Senegal-Akazie (Senegalia senegal)
Gummi arabicum (Harz), Verdickungsmittel,Bindemitte und
Stabilisator in Nahrungsmitteln und Getränken, aber auch in Kosmetikartikel,
Farben u.a. (E 414)
nicht kultiviert
Wüsten- und Halbwüstengebiete südlich der Sahara, v. a.
Senegal
im Nahen Osten wird aus dem Fruchtfleisch Limonade
hergestellt; in Thailand und Mexiko werden kandidierte Früchte als Konfekt gegessen;
die Samen werden gekocht und geröstet und zu Mus gestampft (Tamarindenmark);
gemahlene Samen dienen auch als Geliermittel
Tropen und Subtropen
Afrika
Teparybohne (Phaseolus acutifolius)
junge Hülsen als Gemüse, reife Samen in Suppen
v.a. Mexiko
nördliches Mexiko
Tragant (Astragalus gummifer u.a.Arten)
Harz (Traganth) dient als Verdickungs- und Bindemittel (E
413)
kaum kultiviert
West- und Vorderasien, Iran bis Türkei
Turibaum,
Kolibribaum (Sesbania grandiflora)
Gemüse, Grünfutter, Harze und Gerbstoffe, v. a. für lokale
Nutzung
Schuppenwurz und Gelbe Anemone an der Rotach bei Oberteuringen
Oberteuringen liegt an der Rotach, etwa
in der Mitte ihres knapp 40 km langen Laufes vom Pfrunger-Burgweiler Ried bis
zum Bodensee bei Friedrichhafen. Auf
einem Frühlingsspaziergang entlang dieses Baches lernen wir Bäume, Sträucher
und einige typische Frühlingsblumen kennen, die den Bachlauf begleiten.
Es wird auch gesammelt, sortiert,
gefühlt und geschnuppert.
Samstag, 01.06.2019, 10:00
Uhr
Ersatztermin
bei schlechtem Wetter: 08.06.2019
Treffpunkt: Oberteuringen, Wanderparkplatz Altweiherwiesen (Richtung Bibruck), hinter der Straßen-Unterführung unter der L329
Breitblättriges Knabenkraut, NSG Altweiherwiese bei Oberteuringen
Der zweite Botanische Spaziergang führt uns vorbei an Oberteuringens Naturschutzgebiet Altweiherwiesen mit seinen Feuchtwiesen und Rieden bis zum Weiler Wammeratswatt. An den Wegrändern wachsen viele weitverbreitete Kräuter und Stauden, im Ried hoffen wir auf Knabenkräuter und Sibirische Schwertlilien und einige andere Besonderheiten.
Samstag, 10.08.2019, 10:00 Uhr
muss wegen Krankheit ausfallen
Ersatztermin
bei schlechtem Wetter: 17.8.2019
Treffpunkt: Wanderparkplatz an der Kreisstraße K 7742 zwischen Raderach und Riedheim (am Fuß des Drumlins „Franzenberg“)
Im NSG Hepbach-Leimbacher Ried
Der Spaziergang führt in das Umfeld des Naturschutzgebietes Hepbacher-Leimbacher Ried. Der abwechslungsreiche Weg durch den Wald und vorbei an Feuchtgebieten und Gewässern verspricht nicht nur viele verschiedene Pflanzenarten sondern auch interessante Vogelbeobachtungen.
Samstag, 21.09.2019, 10:00 Uhr
muss wegen Krankheit ausfallen
Treffpunkt: Wanderparkplatz im Brochenzeller Wald an der Landstraße L 329 zwischen Ettenkirch und Brochenzell, von Oberteuringen kommend links, kurz vor dem Kreisverkehr am Ortseingang von Brochenzell
Im Wald bei Brochenzell
Auf diesem Spaziergang gehen wir eine
kleine Runde durch dieses schöne Waldgebiet mit abwechslungsreichem
Baumbestand. Besonders interessieren uns die Pflanzen an den feuchten
Wegrändern und Gräben. Vielleicht finden wir auch einige Pilze.
Dauer jeweils ca. 2,5 Stunden. Die Veranstaltung ist kostenlos.
Auch im Sommersemester 2019 biete ich unter der Veranstaltung „Regionale Lebensräume“ wieder vier ganztägige Exkursionen an. Zwei der Exkursionsziele decken sich mit Angeboten der letzten Jahre:
Neu ist eine Exkursion in das Naturschutzgebiet Altweiherwiese bei Oberteuringen (18.5.2019) und in den Altdorfer Wald mit dem NSG Füremoos bei Vogt (8.6.2019).
Am 24.4.2019 findet von 18:00 Uhr bis 19:15 Uhr eine einführende Informationsveranstaltung in der PH Weingarten, Fach Biologie, NZ 1.51, statt, bei der Erläuterungen zu den Exkursionszielen gegeben und mögliche Aufgaben besprochen werden.
*durch Anklicken kommt man zu Unterlagen für die Exkursion im Sommersemester 2018
Treffpunkt: 10:00h, Wanderparkplatz bei Appenweiler
Thematische Schwerpunkte: Lebensform Baum, ökologische Ansprüche von Waldbäumen und Waldkräutern, Lebenraum Waldgraben
Exkursionsverlauf
Um 10:00 Uhr versammelten wir uns bei leichtem Regen an dem kleinen Wanderparkplatz kurz hinter Appenweiler an dem Sträßchen Richtung Brochenzell. Nach einer Vorstellungsrunde gingen wir auf dem Weg ein kleines Stück am Waldrand entlang und in den Wald hinein. Während des „Bäume Ertastens und Wiedererkennens“ blieb es ziemlich feucht. Wie erwartet wurden die ertasten Bäume – Wald-Kiefern, Linden und Rot-Fichten – relativ leicht wiedergefunden (Anleitung Max Fischer; Markus Preuss). Anschließend – der Nieselregen wurde heftiger – führten wir eine Untersuchung zur Artenzusammensetzung mithilfe der „Wäscheleinen- Transektmethode“ durch.
Wäscheleinen-Tranektmethode
Die Auswertung ergab in dem von uns untersuchten Waldstück eine deutliche Dominanz der Rot-Fichte, gefolgt von der Rot-Buche. Buchen fanden sich vor allem auch im Unterwuchs. Diese Naturverjüngung ist vermutlich von forstlicher Seite beabsichtigt, um auf längere Sicht den Fichtenanteil des Waldes zu verringern (Albijona Sabani).
Nass und kalt
Wetterradar online hatte in Aussicht gestellt, dass
der Regen nach 11:00 Uhr nachlassen würde. Leider regnete es 11:30 Uhr eher
heftiger als vorher und deshalb entschlossen wir uns, etwas Material
einzusammeln, um den zweiten Teil der Exkursion in der pädagogischen Hochschule
Weingarten fortzusetzen. Dort wurden von den Studierenden mit dem
eingesammelten Material Stationen eingerichtet, die dann reihum bearbeitet
wurden.
Wachstum und Alter von Bäumen (mit zwei mitgebrachten Baumscheiben; Anleitung Vanessa Golic, Susanna Wild)
Bäume und Sträucher nach Blättern/Zweigen bestimmen
Ökologische Ansprüche von Wegrandpflanzen und Waldbodenpflanzen vergleichen
Laubstreu untersuchen: Tiere, Zersetzungsstadien der Blätter
Blätterbilder
Kräutermaus aus Stumpfblättrigr Ampfer, Brombeere, Hahnenfuß- und Wiesen-Schaumkraut-Blüte
Zum Schluss wurden folgende Kräuter zur Zubereitung
einer Kräutersuppe genutzt:
Große Brennnessel, Giersch, Gundermann, Kriechender
Günsel, Wiesen-Kerbel, Knoblauchs-Rauke.
Die Kräuter wurden fein geschnitten und in eine
gebundene Suppe eingerührt, die mithilfe von Mehl,, Rapsöl, Wasser und Kräuterbrühwürfeln
zubereitet wurde. Bei der Nutzung von Wildkräutern für die Zubereitung von
Mahlzeiten empfiehlt sich das Abkochen (Suppe, Gemüse, Tee) aus hygienischen
Gründen.
Während die Suppe zubereitet wurde, erhielten wir von Madeleine Mayer Informationen über die Gefahren von Zecken und die Möglichkeiten, sie zu verringern. Hannah Dyx informierte uns über den Lebenszyklus des Fuchsbandwurms und die sich daraus ergebenden Gefahren beim Aufenthalt in der freien Natur.
Treffpunkt: 10:00h, Oberteuringen, Parkmölichkeit bei Unterführung unter L 329 an kleiner Straße Richtung Bibruck; NSG Altweiherwiese
Thematische Schwerpunkte: Verschiedene Wiesentypen insbesondere Streuwiesen, Gräser,Waldrand, Streuobstwiesen, Naturschutz-Pflegemaßnahmen, Landschaftsgeschichte
Das 78 ha große Naturschutzgebiet Altweiherwiesen wurde 1981 vom Regierungspräsidium Tübingen ausgewiesen. Es liegt nordöstlich von Oberteuringen auf einer Meereshöhe von rund 450 m.
Zur Landschaftsgeschichte
Die Niederung ist der Rest einer ehemaligen Schmelzwasserrinne der späten Würmeiszeit (Konstanzer Stadium), die ursprünglich vom Schussenbecken bis ins Salemer Becken reichte und die heute vom Taldorfer Bach durchflossen wird. Ihre Fortsetzung fand sie zuerst im Deggenhauser Tal, das zunächst von der Rotach, hinter Roggenbeuren von der Deggenhauser Aach durchflossen wird. Später bahnten sich die Schmelzwässer den Weg südlich vom Gehrenberg bis zum heutigen Überlinger See.
Verlauf der Schmelzwasserrinne am Ende der Würmkaltzeit vom Schussenbecken durch die Taldorfer Senke (NSG Altweiherwiese) – und weiter südlich entlang der B33 Richtung Markdorf (violett getönt) – Kartengrundlage: Landkreis Ravensburg, Rad- und Wanderkarte 1:50 00 Bodensee (schwarz). Schwarze Linie: Schussen; weiße punktierte Linie: Innere Würmendmoräne (IWEM); weiß gestrichelt: Ausstrich von Endmoräne und lokalen kliffähnlichen Abschürfungsmerkmalen des „Konstanzer Stadiums“ (KS); dünne weiße Linien: Moränenrelikte innerhalb der Endmoräne des Konstanzer Stadiums. Beschriftung von Nord nach Süd und West nach Ost: RAV: Ravensburg, SAL: Salem, ÜBE: Überlingen, GEH: Gehrenberg, MAR: Markdorf, KON: Konstanz, FRI: Friedrichshafen, TET: Tettnang. 1-3 verschiedene Drumlins. Die Schmelzwasserrinne, die das Schussensbecken mit dem Salemer Becken verbindet. ist gut zu erkennen.
Quelle: Elena Beckenbach, Thomas Müller, Hartmut Seyfried, Theo Simon (2014): Potential of a high-resolution dtm with large spatial coverage for visualization, identification and interpretation of young (Würmian) glacial geomorphology a case study from Oberschwaben (southern Germany) Quaternary Science Journal Volume 63 / number 2 / 2014 / 107–129 / DOi 10.3285/eg.63.2.01
Im Bereich des NSG trennen tonige Sedmente den Untergrund zu den eiszeitlichen Schottern hin ab. Durch den Wasserstau kam es am Ende der Würmeiszeit zur Moorbildung. Die heutige L 329 verläuft auf einem im Mittelalter durch Mönche des Klosters St. Gallen errichteten Damm, der die Moorfläche zu einem großen Fischweiher werden ließ. Im 18 JH wurde das Wasser wieder abgelassen, um die Sumpfbereiche als Streuwiesen nutzen zu können. Bis Mitte des vorigen Jahrhunderts wurden das Mähgut von den umliegenden Bauernhöfer als Einstreu verwendet. Heute werden die Schilfflächen regelmäßig vom Naturschutz gemäht, um die Streuwiesen-typische Vegetation zu erhalten.
Flora, Fauna, Lebensräume
Wichtigster schützenswerter Lebensraum sind magere Flachlandmähwesen und Gewässersäume mit Erlen und Weiden. Dort gedeihen große Bestände des Breitblättrigen Wollgrases (Eriophorum latifolium), außerdem Pracht-Nelke (Dianthus superbus), Böhmischer Beinwell (Symphytum officinale ssp.bohemicum), Sibirische Schwertlilie (Iris sibirica), Berg-Klee (Trifolium montanum), Breitblättriges und Fleischfarbenes Knabenkraut (Dactylorhiza majalis und D. incarnata) und Mücken-Händelwurz (Gymnadenia conopsea).
Würden die Wiesen nicht regelmäßig gemäht, würden sich daraus schnell reine Schilfbestände entwickeln.
An Greifvögeln kann man regelmäßig Bussarde, Turmfalken und Rote und Schwarze Milane beobachten. den für das Gebiet angegebenen Baumfalken konnte ich allerdings noch nie sehen.Weitere Besonderheiten sind Kleinspecht, Nachtigall, Schwanzmeisen und Neuntöter. Im Taldorfer Bach leben Elritzen, verschiedene Weißfische und auch Hechte. Oft kann man in Bachnähe auch Ringelnattern entdecken.
Naturschutzmanagement
Im Jahr 2017 wurde aufgrund einer Kartierung für die Rotach einschließlich des Naturschutzgebietes Altweiherwiese (FFH-Gebiet 8222 – 342) ein Managementplan erstellt. Die für das Naturschutzgebiet vorgesehenen Pflegemaßnahmen sind in einer Karte dargestellt. Dabei bedeuten die mich violetter Farbe gekennzeichneten Flächen, dass auf diesen eine einschürige Mahd vorgesehen ist.
Regierungspräsidium Tübingen (Hrsg.): Managementplan für das FFH-Gebiet 8222-342 „Rotachtal Bodensee“ . bearbeitet von INULA, Freiburg. 1. Dezember 2017 (174 S., baden-wuerttemberg.de [PDF]).
Ein gewisses Problem für das Naturschutzgebiet stellt der Kraftfahrzeugverkehr auf dem Sträßchen dar, das über Bibruck, Reute, Sederlitz bis nach Dürnast führt und dort in die B 33 mündet. Die Straße wird leider nicht nur von Anwohnern sondern auch als Shortcut von der K 329 zur B 33 und umgekehrt genutzt. Neben verkehrsberuhigten Maßnahmen („nur für Anlieger“) wäre auch an einen Bohlenweg parallel zum Sträßchen zu denken, der von Fußgängern (und Radfahrern?) genutzt werden könnte.
Wammeratswatt
Am Rande des Naturschutzgebietes liegt ein Weiler mit dem eigentümlichen Namen Wammeratswatt. Ein altes zwischen Vegetation verstecktes Schild weist darauf hin, dass dieser Ort bereits im zwölften Jahrhundert, 1164, als Wambrehteswathe urkundlich erwähnt wurde. Die Endung “-wathe” geht wohl auf einen altgermanischen Begriff zurück, der so viel wie “Weide” oder “Wiese” bedeutet. “Wanbrecht” ist ein alemannischer Name. Man könnte die Ortsbezeichnung also mit „Weide oder Wiese des Wanbrecht“ übersetzen.
Foto Uwe Baur, 23.6.2012
Exkursionsverlauf
Exkursionsweg am 18.5.2019; Ausschnitt aus TK 8222,Markdorf, 1:25000
Bei trockenem Wetter und angenehmen Temperaturen treffen wir
uns um 10:00 Uhr in Oberteuringen an der Unterführung unter der Landstraße 329 am
Eingang zu dem Naturschutzgebietes Altweiherwiese.
Nach einer Einführung in die Landschaftsgeschichte und die
Besonderheiten des Naturschutzgebietes beschäftigen wir uns mit dem Stockwerkaufbau
einer Wiese.
Auf dem Weg Richtung Wammeratswatt werden zunächst möglichst
viele unterschiedliche Gräser gesammelt, sortiert und bestimmt. Anschließend
versuchen wir, die verschiedenen Grasarten allein aufgrund haotischer Merkmale,
also mit verbundenen Augen, zu erkennen. In dem langsam fließenden Taldorfer
Bach wachsen dichte Bestände des Wassersterns, von denen dank des Einsatzes von
Markus Preuss alle TeilnehmerInnen auch einen Zweig in den Händen halten
können.
Aufgrund der vorangegangenen relativ kühlen Witterung ist die
Vegetationsentwicklung auf der Streuwiese noch ziemlich weit zurück – keine Sibirischen
Schwertlilien, Pracht-Nelken oder fruchtenden Wollgräser sind zu sehen. Von den
charakteristischen Arten können wir nur gerade aufblühende Exemplare des
Breitblättrigen und des Fleischfarbenen Knabenkrautes entdecken, außerdem Berg-Klee,
Sumpf-Kratzdistel, Gewöhnlichen Hornklee und Gilbweiderich (noch nicht blühend).
Auf dem weiteren Weg zum Weiler Wammeratswatt fallen die als Straßenbegleitbäume gepflanzten Robinien auf, die gerade erst ihre Fiederblätter entfalten. Ebenso wie die alten Obstbäume der Streuobstwiese sind sie reichlich mit Misteln bewachsen. Die Bäume der Streuobstwiese – vorwiegend Äpfel auch einige Birnen – sind mehr als 100 Jahre alt. Sie werden immer noch zur Gewinnung von Fallobst für die Apfelsaftproduktion genutzt, das Heu der Wiese wird an Reiterhöfe verkauft. In der Wiese fällt uns der Kleine Wiesenknopf mit seinen weit aus den Blüten heraushängenden Staubfäden besonders auf.
Kleiner Wiesenknopf – Sanguisorba minor
Nach einer Mittagspause bei der Brücke über den Taldorfer Bach
leiten Yesim Örgerim und Beatrice Hell
zu „Blüten, lockende Signale“ an. Blütenteile – die Bestäuber anlocken sollen –
und Blätter werden zu Make-up oder schmückenden Kurzzeit-Tattoos umgewidmet.
Der Weg führt uns dann weiter dem Waldrand entlang. Unter Anleitung von Lisa-Marie Buemann und Sara Dittmann sammeln wir Tierspuren: Spuren an Blättern, Spuren an Zapfen und Früchten und Spuren an Holz. Besonders auffällig sind die Minen des Buchen-Springrüsslers an Rot-Buchen-Bättern.
Hier hat ein Specht einen Fichtenzapfen bearbeitet
Unser Exkursionsweg führt dann weiter zum Weiler
Blankenried. Dort kreuzen wir die L 329 und besteigen den Drumlin Horach (501,6
m ü.N.N.), auf dem sich ein Wasserreservoir der Wasserversorgung Bodensee
befindet. Von dort hat man eine sehr schöne Aussicht auf den Bodensee und die
Alpenkette – heute allerdings im Dunst verborgen. Sabrina Brendle führt die
Gruppe in einer blinden Raupe aus dem Wald heraus zum Aussichtsplatz, wo die
Augenbinde dann abgenommen werden.
Altdorfer Wald mit NSG Füremoos
Gelb umrandeter Ausschnitt unten:
Treffpunkt: 10:00h, Wanderparkplatz am Waldrand bei Vogt
Der Altdorfer Wald liegt auf einem Höhenzug zwischen Vogt im Süden und auch Aulendorf im Norden am nordöstlichen Rand des Schussenbeckens. Mit 82 km2 ist er das größte zusammenhängende Waldgebiet Oberschwabens. Der Name geht auf die welfische Grafschaft Altdorf bzw. auf die ehemalige Ortsbezeichnung „Altdorf“ für die Stadt Weingarten zurück. Bis 1865 wurde lediglich das Kloster als „Weingarten“ bezeichnet, während die umgebende Ortschaft Altdorf genannt wurde.
Naturraum
Der Altdorfer Wald besteht hauptsächlich aus Fichtenforsten, eingesprengt sind aber auch Buchen und andere Laubbäume. Er enthält die Naturschutzgebiete Saßweiher, Girasmoos, Tuffsteinbruch Weißenbronnen, Lochmoos und Füremoossowie ein Fauna-Flora-Habitat Gebiet „Altdorfer Wald“ mit 13,5 km2. Der Höhenzug übersteigt an einigen Stellen700 m, höchste Erhebung ist der Galgenberg mit 776,6 m ü.N.N.
Bedeutendstes Fließgewässer ist die Wolfegger Ach, die wie einige weitere kleinere Zuflüsse in die Schützen mündet und einige weitere kleinere Bäche der Schussen zufließt. In dem Waldgebiet liegen auch einige Seen, wie der Bunkhofer Weiher, der Neuweiher und der Langmoosweiher und aus einigen feuchten Senken haben sich Moore entwickelt. Eines davon, das Naturschutzgebiet Füremoos, werden uns genauer anschauen. (Weitere Unterlagen zum Thema Moore und Feuchtgebiete in den Unterlagen zur Exkursion ins Wurzacher Ried 2018):
Die Kalktuffbildungen bei Weißenbronnen weisen auf kalkreiches Jungmoränenmaterial im Untergrund hin.
Thematische Schwerpunkte: Konzeption des Naturschutzzentrums Wilhelmsdorf, Landschaftsgeschichte, Insekten
Das digitale Geländemodell zeigt, wie sich das Pfrunger Becken zwischen der äußeren un inneren Endmoräne der Würm-Kaltzeit ausgebildet hat.
Zum Exkursionsverlauf
Nach der Begrüßung durch Frau Ackermann, Diplom-Biologin und
Naturpädagogin am Naturschutzzentrums Wilhelmsdorf, beschlossen wir aufgrund
der Wetterlage, die für den Nachmittag geplante Exkursion zum Fünfeckweiher und
zum Bannwaldturm schon am Vormittag durchzuführen.
Startpunkt war – wie in den vergangenen Jahren -der
Parkplatz bei Uzhausen. An einer Übersichtskarte erklärte uns Frau Ackermann
die verschiedenen Schutzzonen des Pfrunger-Burgweiler Rieds und die jeweiligen Schutzziele
und -maßnahmen. Das Pfrunger-Burgweiler
Ried ist nach dem Federseegebiet das zweitgrößte zusammenhängende Moorgebiet
Südwestdeutschlands, die größte zusammenhängende Hochmoorfläche allerdings ist
im Wurzacher Ried zu finden.
Zunächst ging unser Weg durch Weideland, auf dem wir eine
Herde Scottish Highlander beobachten konnten, die hier zusammen mit eigen
anderen Robustrinderrassen zur Biotoppflege eingesetzt werden. Sie sind das
ganze Jahr über auf der Weide, werden allerdings im Winter mit Heu zugefüttert,
das im Sommer auf den Weideflächen gewonnen wird. Fleisch und Wurst waren von
den Robustrinder sind im Naturschutzzentrum Wilhelmsdorf erhältlich, außerdem
werden sie in einigen Gaststätten rund um das Pfrunger-Burgweiler Ried
angeboten.
Entlang eines ehemaligen Entwässerungskanals, der in
regelmäßigen Abständen durch Querwände aufgestaut ist, konnten wir auf
Weidezäunen mehrfach Schwarzkehlchen und einen Neuntöter beobachten. Im
Feuchtbereich um den Graben blühte reichlich Mädesüß. Auf den Dolden des
Wiesen-Bärenklaus waren viele Nektar sammelnde Insekten, insbesondere Schwebfliegen
und kleine Bockkäfer, zu beobachten. Am Wegrand nahmen wir eine Geruchsprobe
vom Feld-Thymian, der gerade in voller Blüte stand.
Auf den Weideflächen brüten regelmäßig Kiebitze, für die in
diesem Jahr extra flache Gewässer, „Blänken“, angelegt wurden, die vor allem
für die jungen Kiebitze wichtig sind, weil sie dort leicht an geeignete Nahrung
(Insektenlarven, Würmer) kommen können.
Wir folgten dann dem Weg durch den Bannwald bis zum Fünfeckweiher.
In Baden-Württemberg bezeichnet der Begriff „Bannwald“ Totalschutzgebiete, die
vollständig einer natürlichen Entwicklung überlassen werden. Wege durch den
Bannwald sind zulässig, und dürfen – zum Beispiel durch Fällen umsturzgefährdeter
Bäume – gesichert werden. Auf dem Weg fällt auf, dass durch Wiedervernässung
die Fichtenbestände links des Weges großflächig abgestorben sind. Den Fünfeckweiher
erreichten wir auf einem Bohlenweg. Wie in den Vorjahren konnten wir eine ganze
Reihe von Insekten beobachten, insbesondere die dunkle Sommergeneration des
Landkärtchens, eines Schmetterlings, der oft auch in Schulbüchern als Beispiel
für die Ausbildung unterschiedlich aussehender Generationen angeführt wird.
Von der Aussichtsplattform des Bannwaldturmes hatten wir
einen guten Überblick über das gesamte Gebiet des Pfrunger-Burgweiler Rieds und
die umgebenden Höhenzüge. Man kann von dort aus sehr gut die verschiedenen
durch industriellen Torfstich entstandenen Seen beobachten. Eine kleine Schar
Graugänse flog am Turm vorbei.
Am sonnigen Wegrand wurde auf zwei Stammformen von
Kulturpflanzen hingewiesen, die hier besonders schön entwickelt waren: Wilde
Möhre (Daucus carota) und Wegwarte oder Zichorie (Cichorium intybus), aus der sowohl der Chicoréesalat als auch die
Wurzelzichorie (Zichorienkaffee) gezüchtet wurde. Bei unserem Vesperplatz konnten
wir ein schönes Exemplar des Kompass-Lattichs oder Stachel-Lattichs (Lactuca serriola) anschauen. Sein Name
rührt daher, dass sich seine Blätter an sonnigen Standorten senkrecht stellen
und in Nord-Süd-Richtung orientieren. Damit sind die Blattspreitenen weniger
dem intensiven Sonnenlicht ausgesetzt. Der Kompass-Lattich ist die Stammform des
Gartensalates (Lactuca sativa).
Da um uns herum erhebliche Gewitteraktivitäten zu beobachten
waren und die Zeit auch schon recht fortgeschritten war, beschlossen wir, auf
einen größeren Rundweg (vergleiche Exkursionen der Vorjahre) zu verzichten und
direkt zum Parkplatz bei Holzhausen zurückzukehren. Auf den Weiden in der in
der Nähe des Parkplatzes konnten wir eine große Zahl Weißstörche (gezählt
wurden 36) beobachten.
Da es bei unserer Ankunft am Naturschutzzentrum Wilhelmsdorf
immer noch relativ gutes Wetter war, beschlossen wir, nun gleich mit dem Fang
von Wiesentieren und Wassertieren zu beginnen. Ausgerüstet mit Keschern und
Wasserschalen bzw. Schmetterlingsnetzen und Becherlupen machten sich zwei
Gruppen zunächst auf die Jagd, dann würden die Tiere im Großraum mithilfe von
Bestimmungsschlüsseln und Binokularen untersucht und bestimmt. Einige Fänger
konnten über den Monitor demonstriert werden.
Einige unserer Fänge:
Wiese
Larve einer Kurzfühler-Heuschrecke (vielleicht Corthippus
parallelus)
Schwebfliege
Mücke mit gelbem Hinterleib
Baumwanze
Weichwanze
Schaumzikade
Brauner Waldvogel
verschiedene Kleinschmetterling (Zünsler?).
Steinhummel
Teich
Wassermilbe
Kleinlibellenlarve
Stabwanze
Rückenschwimmer
Schwimmwanze
Büschelmückenlarve
Käferlarve
Kaulquappe eines Wasserfrosches (Kleiner Teichfrosch?)
Beim abschließenden Museumsbesuch gibt uns Frau Ackermann anhand eines großen Luftbilds der Region einen Überblick über die Entstehung des Pfrunger-Burgweiler Rieds. Auf der Fahrt in dem „Moorkäpsele“ werden diese Fachinhalte anschaulich wiederholt (vergleiche Exkursionsbericht von 2017).
Mögliche Aktivitäten von Studierenden
Anleitungen
Baum ertasten und wiedererkennen
Wachstum und Alter von Bäumen
Bäume zählen
Bäume berechnen
Baumkronenspaziergang
Kräuter fühlen, riechen, schmecken
Gräserberührungen
Pflanzenoberflächen: Rau und glatt und andere Gegensätze
Blüten, lockende Signale
Vertauschte Gegenstände
Wer war der Übeltäter?
Umwelt im Umschlag
Torfmoose und Moorbildung
Über alle Exkursionen:
Artenliste Pflanzen
Artenliste Tiere
Gut erkennbare Pflanzenfamilien besonders beachten:
Nach der Blutmahlzeit suchen Zeckenweibchen eine geschützte Stelle und legen mehrere 1000 Eier ab, aus denen zunächst sechsbeinige Larven schlüpfen, danach sterben sie (im Bild vermutlich Ixodes ricinus, der Holzbock; Foto: Kämmer 2019).
Fuchsbandwurm
Literaturhinweise
Einen sehr guten Einstieg in die Landschaftsgeschichte Oberschwabens und ganz Südwestdeutschlands kann man sich mit folgendem neu erschienenen Werk verschaffen:
Seyfried, H., Simon, T., Beckenbach, E. & Müller, T. (2019): Der Südwesten im digitalen Geländemodell – wie LiDAR-Daten unsere Sicht auf die Welt verändern. Sonderbände der Gesellschaft für Naturkunde in Württemberg, 4; 434 S., 301 Abb. – Schmidt-Verlag. 34,90 €
















































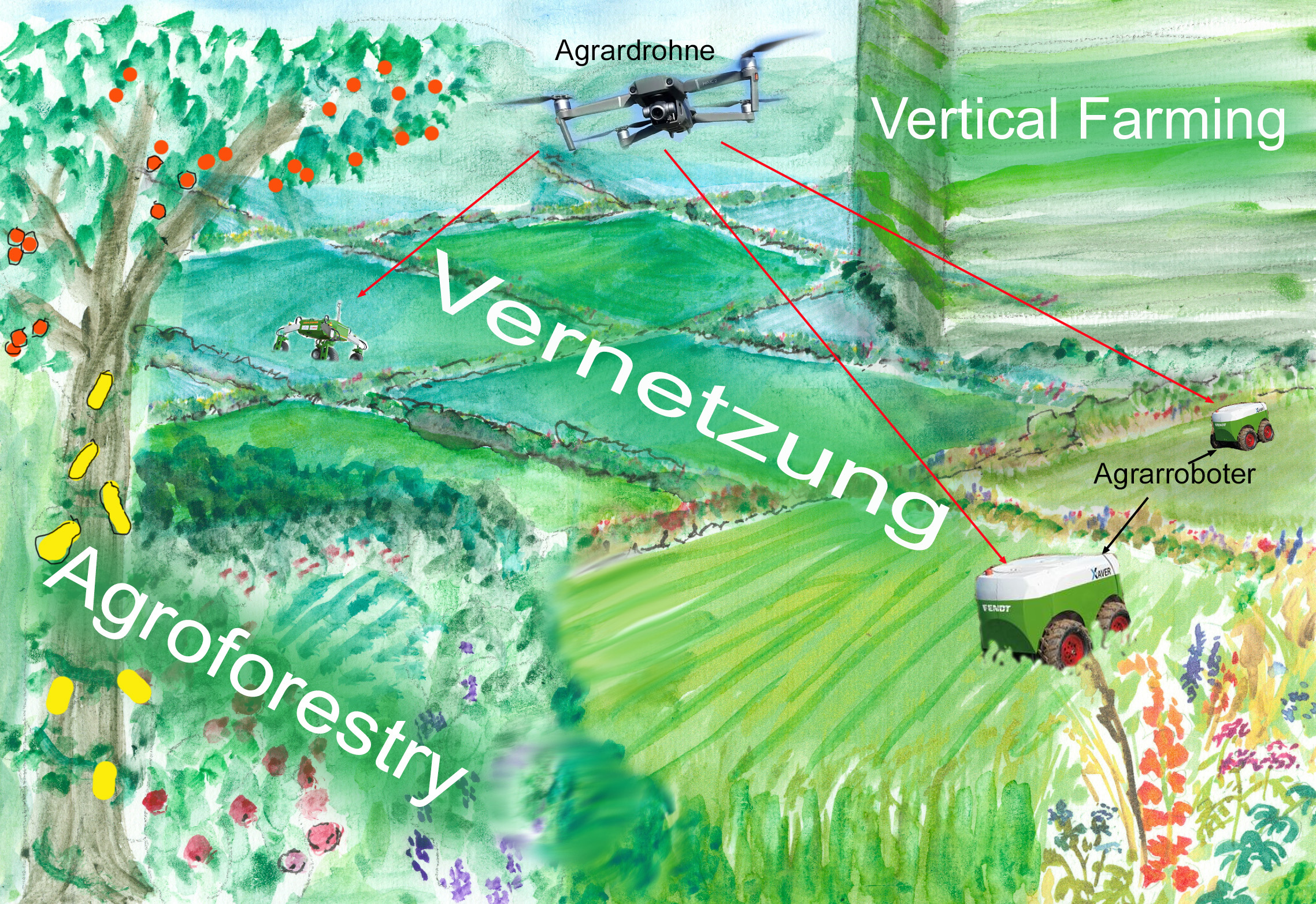

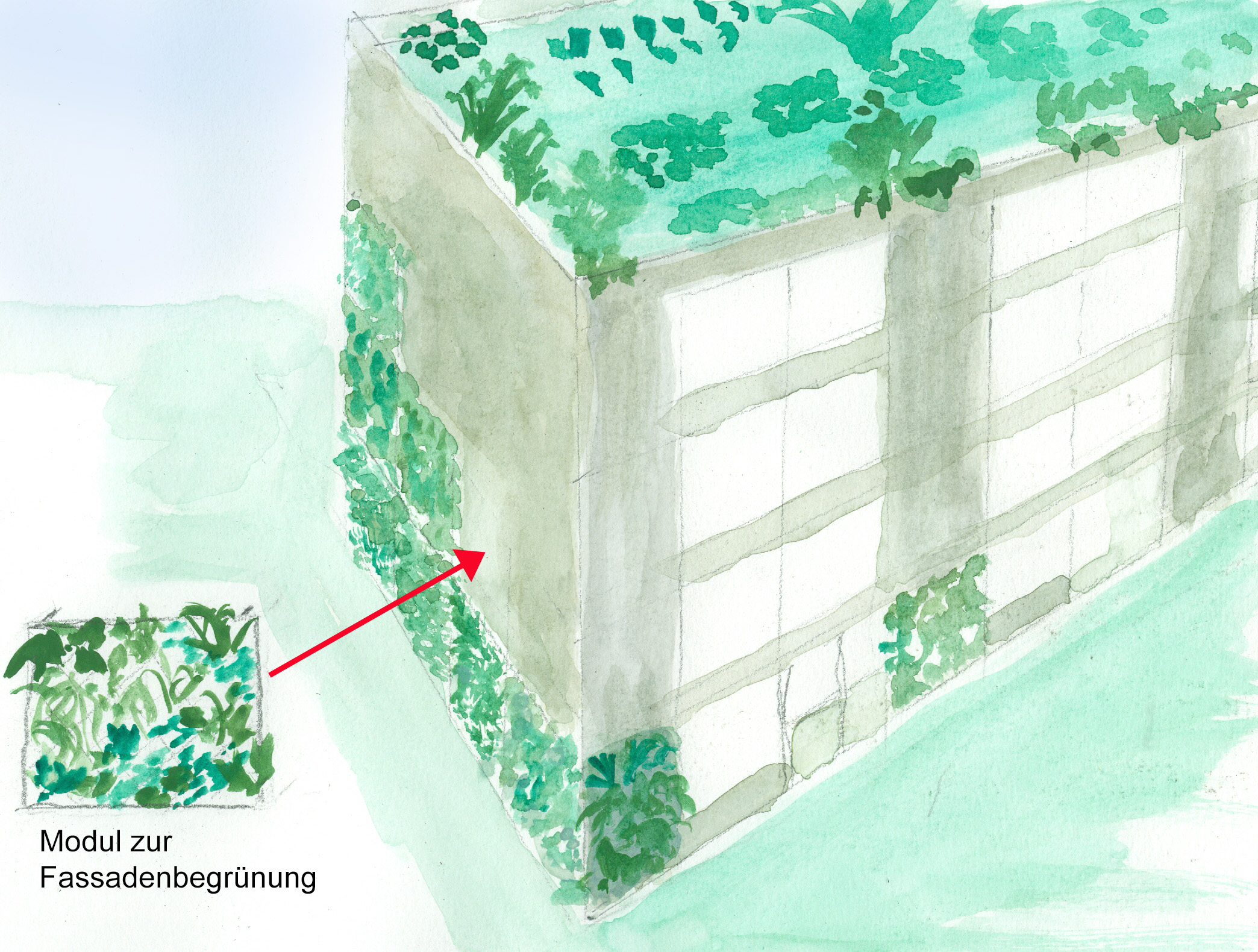










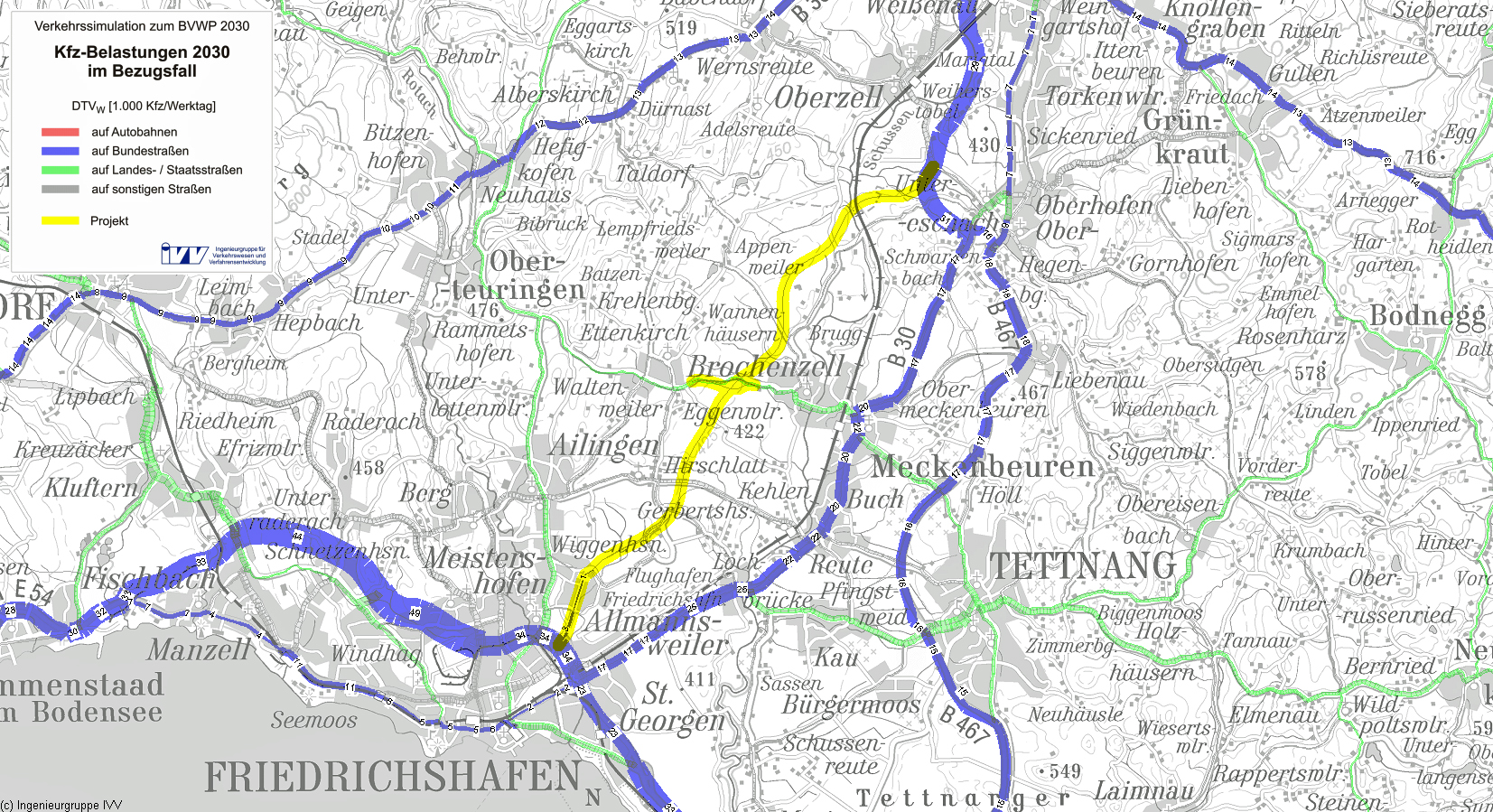





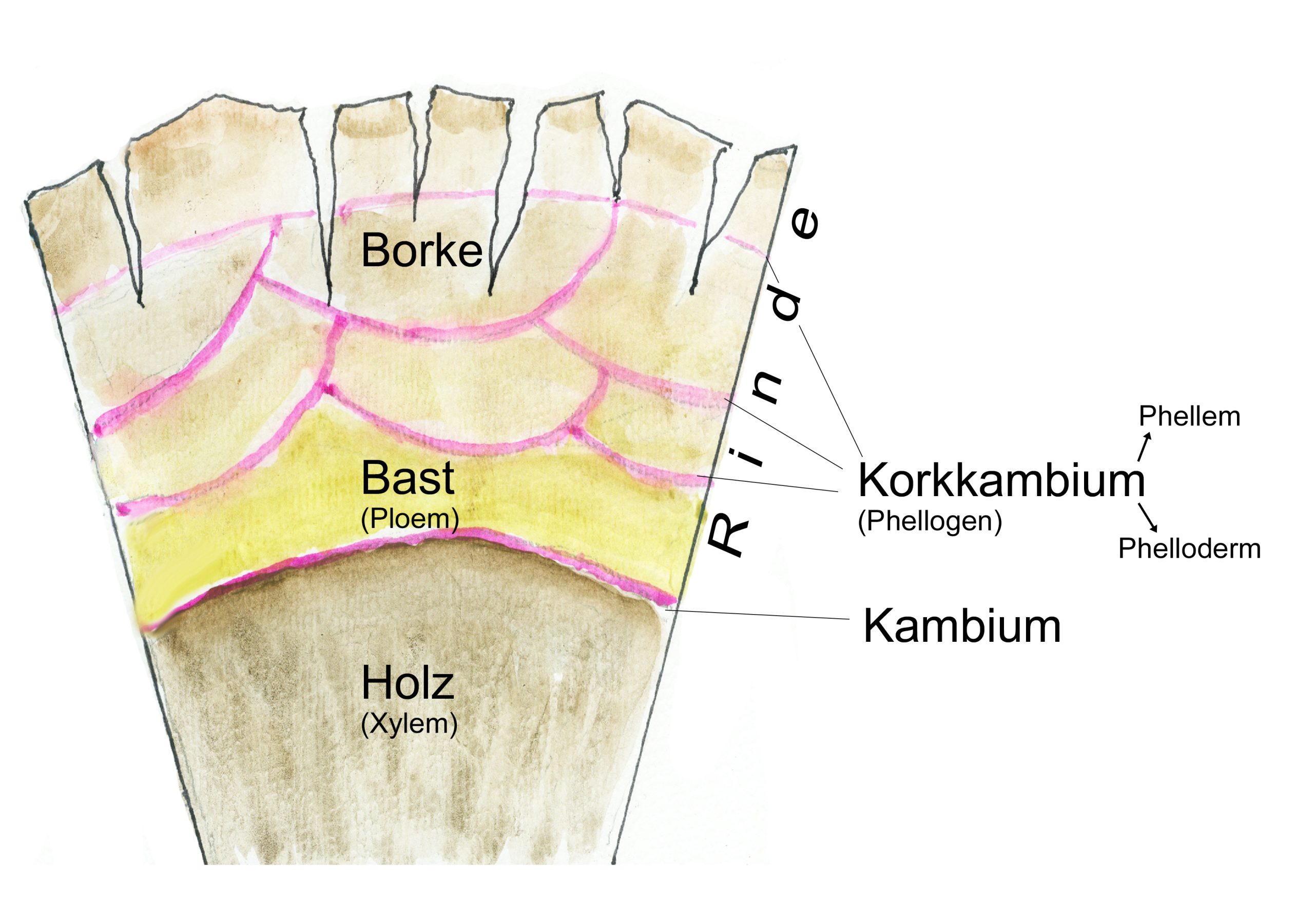









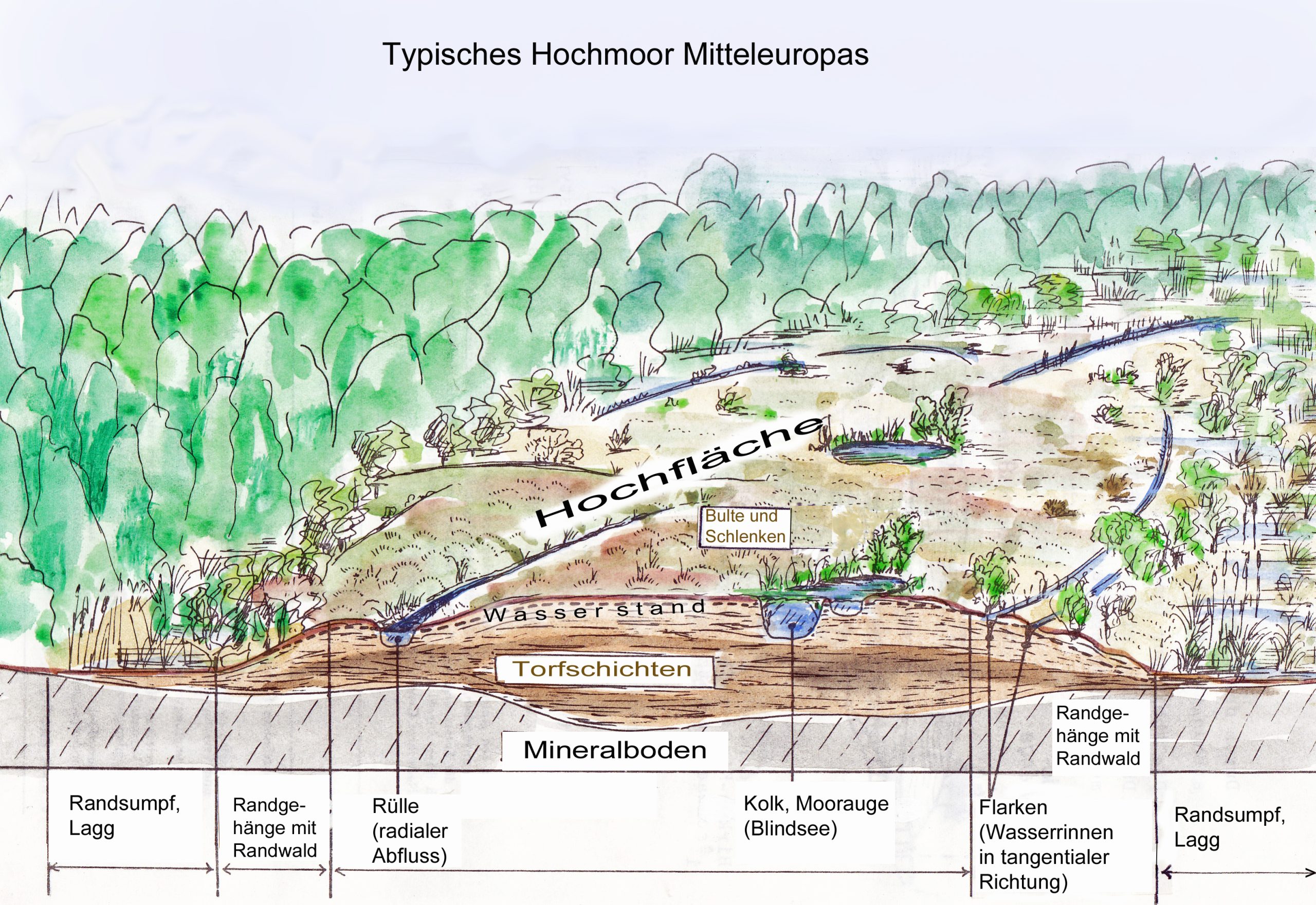









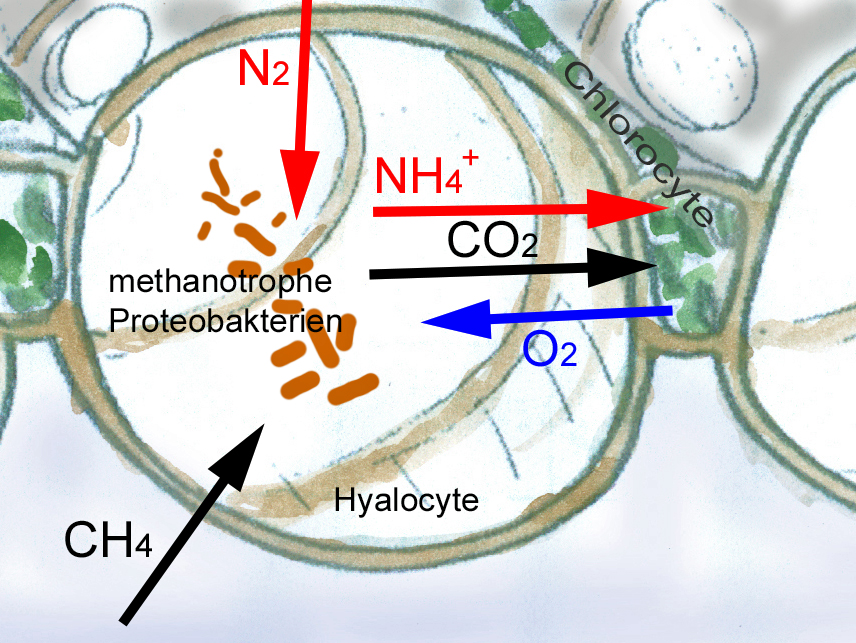



































































































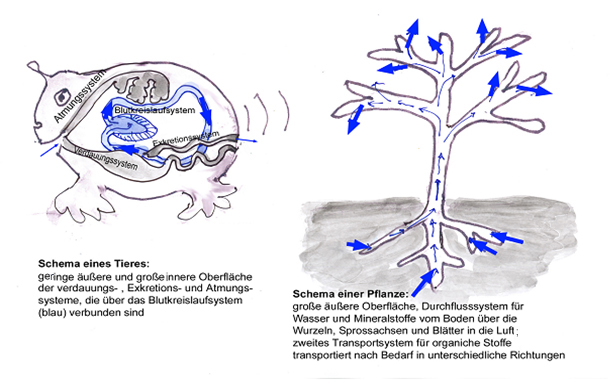




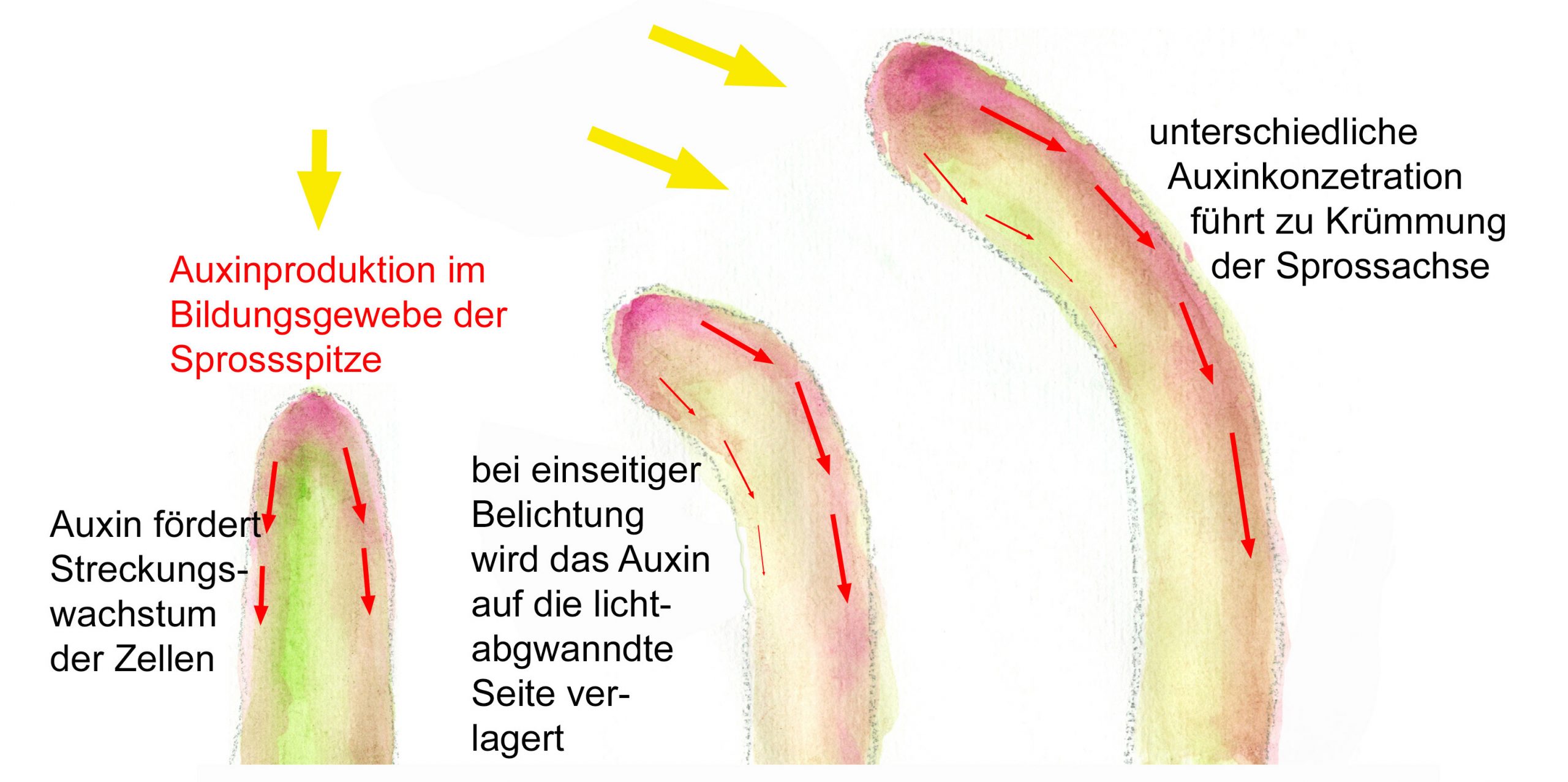






























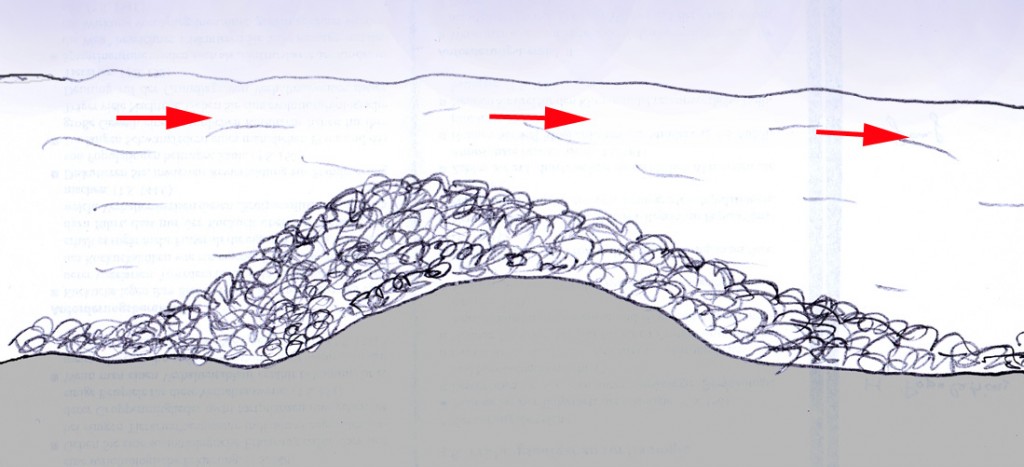



























































.jpg){kind=link}
{kind=link}
{kind=link}